Research 研究成果の紹介
植物は葉の病原菌感染を防ぐ細胞壁強化酵素を根で栄養獲得にも活用していた必須元素のリンの吸収力を向上
植物の免疫制御因子の新たな働きを発見
植物は葉の病原菌感染を防ぐ細胞壁強化酵素を根で栄養獲得にも活用していた
~必須元素リンの吸収力を向上~
植物の免疫制御因子の新たな働きを発見
【概要】
奈良先端科学技術大学院大学(学長:塩﨑一裕)先端科学技術研究科 バイオサイエンス領域の西條雄介教授らの研究グループは、名古屋大学、北海道大学、ケルン大学(ドイツ)との共同研究を通して、植物の根毛(注1)において病原菌抵抗性を誘導する免疫制御因子が、必須元素であるリン(P)を栄養として獲得する際にも重要な働きをしていることを明らかにしました。
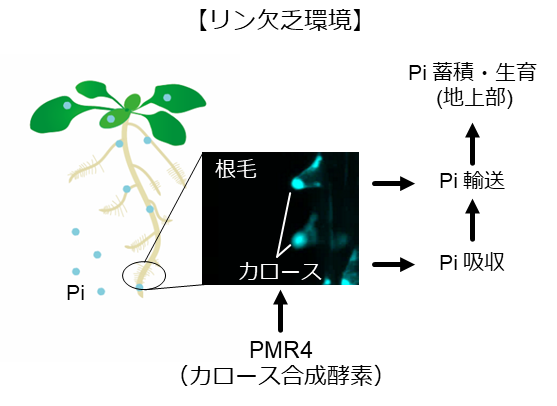
植物は、リンを根から吸収可能な水溶性の無機態リン酸(Pi)の形で土壌から獲得します。しかしながら、リンは不溶化しやすいので、植物が利用可能な Pi は土壌中で不足しがちです。そこで、リン栄養が欠乏した環境では、植物は Pi の吸収・輸送効率を向上させるために一連の生理的・形態的な変化(リン枯渇応答(注 2): PSR と総称される)を誘導します。土壌表層に不溶化されトラップされていることが多いリンを求める手段として、主根を土中深く伸長するのを止めて、その代わりに細い側根や根毛を発達させて横方向に根を広げることが知られています。その際、根毛において、カロース(注3)と呼ばれる多糖類が細胞膜上に蓄積して細胞壁が厚化されることを岡田博士・西條教授らが見出し、その生物学的な役割や制御因子について解明を進めました。
実験では、まずモデル植物のシロイヌナズナを用い、無菌のリン欠乏条件において根毛にカロースを蓄積できない変異体を得ました。次いで、その変異の原因遺伝子が真菌(カビ)病抵抗性に重要なカロース合成酵素遺伝子 GSL5 (PMR4)であること、したがって PMR4 が根毛でのカロース蓄積に必要であることを突き止めました。
さらに、既知の主要な PSR 制御因子の欠損変異体では根毛にカロースが正常に蓄積されますが、その一方で、PMR4 欠損変異体ではカロース欠損が Pi の吸収や地上部への輸送の低下と結びついていることを示しました。つまり、PMR4 は既知の PSR とは異なる様式でリン栄養の獲得に働いていたのです。また、根毛だけに限定して PMR4 を発現させるだけでもリン欠乏環境において地上部の生育を回復させるには十分であることから、免疫制御因子が貧栄養環境への適応にも働いていることを明らかにしました(図 1)。
このような研究成果から、シロイヌナズナが持つ 12 種類のカロース合成酵素遺伝子の中で、PMR4が病害抵抗性にもリン栄養獲得にも寄与していることがわかり、植物の免疫応答と栄養応答が密接な関係性にあることが浮き彫りになりました。
本研究成果は、米国の植物科学雑誌「The Plant Journal」に 2024 年 11 月 15 日(金)午後 5 時 30 分(日本時間)に公開されます(DOI:10.1111/tpj.17134)。
【背景と目的】
植物は、リン栄養欠乏環境では、根からの Pi 吸収や体内での Pi 利用効率を高めるためのリン枯渇応答(PSR)を誘導します。これまで、シロイヌナズナなどを用いた研究から、転写制御因子(注 4) のPHR1・PHL1 や分泌性フェロキシダーゼ(注 5)のLPR1・LPR2 が植物のリン枯渇環境への適応において重要な働きをすることが知られています。PSR に伴い様々な形態変化や生理変化が誘導されますが、その役割や制御因子に関して依然として解明されていない点が多くあります。その一つとして、岡田博士・西條教授らは、根毛においてカロース(β-1,3-グルカンからなる多糖類)の蓄積が誘導されることを見出しました。カロースは、植物の発生・生育の様々な過程において細胞壁と細胞膜の間に蓄積し、例えば病原真菌の葉への侵入に対する障壁や細胞間のシグナル分子の輸送調整などにおいて重要な役割を担うことが知られています。しかし、根の固着や土壌からの栄養吸収に重要な根毛において、リン枯渇環境で誘導されるカロースの役割は不明でした。そのため、根毛でのカロース蓄積の制御因子を同定し、根毛にカロースを蓄積できなくなった変異体でどのような異常が生じるかを解析することで、上記の問題の解明を進めました。
【研究の内容】
低リン条件において、シロイヌナズナのPHR1・PHL1 及び LPR1・LPR2 を同時に欠損した変異体の根毛には正常にカロース蓄積が誘導されることが分かりました。よって、根毛カロース蓄積は、これらの PSR 制御因子から独立して働く新規の制御因子に依存していると考えられました。そこで、EMS 変異原処理(注 6)によって得られたシロイヌナズナ変異体集団から根毛カロース蓄積が低下した caps変異体をスクリーニングしました。得られた変異体の原因遺伝子が、病原真菌に対する侵入抵抗性や免疫応答の際のカロース蓄積に必要なカロース合成酵素遺伝子 GSL5(PMR4)であることを突き止めました(図 2)。
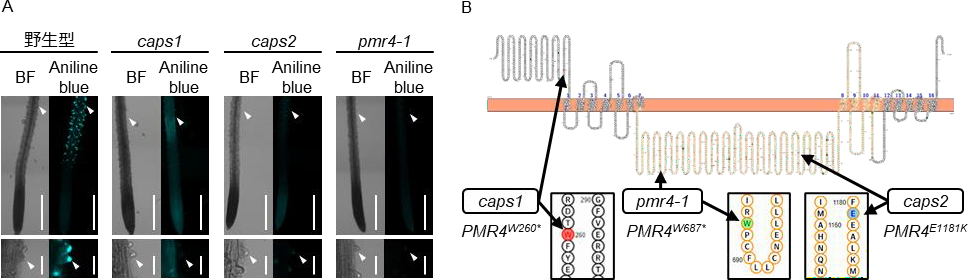
A. EMS 変異体集団から選抜された変異体(caps)と pmr4-1 変異体のカロース蓄積の観察
BF: 明視野、Aniline blue: カロースを検出するための染色。白矢頭は根毛を指す
B. 変異体の PMR4 ペプチド配列における変異箇所
pmr4 変異体では、PSR に特徴的な根毛の伸長や主根の伸長抑制が野生型より顕著に起こっており、PMR4 がリン枯渇環境における根の形態変化にも働いていることが示唆されました(図 3A)。また、pmr4変異体では、リン枯渇状態の植物で引き起こされる生育の低下やアントシアニンの過剰蓄積も強く誘導されており、PMR4 は、PSR を促進するように働くことが示唆されました(図 3B)。こうした PSR に伴う生理変化の大部分は、PHR1・PHL1 やLPR1・LPR2 に依存していることが知られています。PMR4 は、PHR1・PHL1 や LPR1・LPR2 に依存せずに根毛カロースを誘導し、PHR1・PHL1 や LPR1・LPR2 を介した応答が過度に活性化され過ぎないように調節する働きを持つと考えられました。次に、植物体内の Pi 濃度測定試験や 32P(注 7)という放射性同位体を用いて Pi の取り込みを追跡した試験により、Pi の体内濃度や吸収速度、及び根から地上部への輸送効率がpmr4 変異体では低下していることが示され、PMR4 は、Pi の吸収及び輸送に寄与することがわかりました(図 3C)。
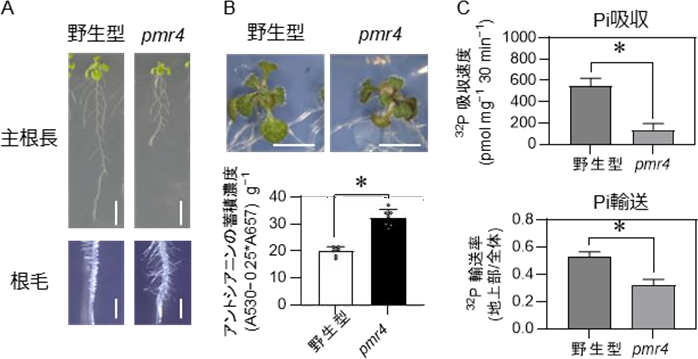
A.主根と根毛 B. 地上部の生育とアントシアニンの蓄積濃度
C.32P を用いた取り込み試験による 32P の吸収速度と輸送率
根または根毛でPMR4 が働くことが上記のPMR4 機能に重要であるかを地上部と根の接木植物(注 8) や、根毛特異的に PMR4 を発現させた植物を作出して調べました。野生型と pmr4 変異体を接木した植物を解析することで、根の PMR4 が地上部の生育に寄与することを示しました。さらに、PMR4 を根毛だけに限定して発現させた場合、地上部の生育は正常に回復されたことから、根毛の PMR4 がリン欠乏環境への適応に働くことが確認されました(図 4)。

A. 根毛特異的に PMR4 を発現させた系統(pLRX1::PMR4/pmr4)のカロース蓄積
B. 根毛特異的に PMR4 を発現させた系統の地上部生重量
【今後の展開】
本研究は、免疫応答時のカロース蓄積に働く PMR4 が、根毛でのカロース蓄積を介してリン栄養獲得に働くことを示すことで、植物の免疫制御と栄養応答調節に密接な関係性があることを強調する発見となりました。根は、栄養吸収に加えて土壌微生物との相互作用の場でもあります。PMR4 をはじめとした免疫制御因子が栄養応答にも働くことで、両応答の密接な連携が可能になり、例えば、土壌や体内の栄養状態に応じて土壌由来の微生物との共生が柔軟に調節できるようになると考えられます。さらに、岡田博士・西條教授らは、本研究に先立ち、植物の免疫を活性化させる Pep ペプチドが根毛形成も誘導する働きを持つこと、及び Pep ペプチドの受容体を根毛だけに限定して発現させた場合も根全体に防御応答を正常に誘導できることを示しています(Okada et al., New Phytologist, 2021)。両研究を通じて、根毛において免疫応答と栄養応答が連携されている様子が示され、今後さらにその仕組みの解明を進めることで植物の栄養欠乏土壌への適応戦略に関する理解も深まるものと期待されます。
【用語解説】
注 1 根毛:植物の根の表皮細胞が外側に向かって管状に伸びた組織。
注 2 リン枯渇応答:無機態リン酸の吸収・輸送効率を向上させる適応応答。主根の伸長を抑制して側根・根毛の伸長を促進するなど根の形態変化、及びリン酸トランスポーターの発現上昇等が誘導される。
注 3 カロース:β-1, 3 グルカンからなる多糖であり、細胞壁、細胞板、原形質連絡に局所的に蓄積し、物理的また生理学的に障壁として働く。
注 4 転写制御因子:DNA に直接結合して、遺伝子の転写(DNA を鋳型にして RNA を複製)促進または抑制するタンパク質。
注 5 分泌性フェロキシダーゼ:第二鉄を第三鉄に酸化する細胞壁局在の酸化酵素。
注 6 EMS 変異原処理:メタンスルホン酸エチルにより種子を変異原処理して変異体集団を得る方法。
注 7 32P: リン(P)の放射性同位体であり、P の動態を追跡するのに使用される。
注 8 接木植物:異なる 2 つの植物をつなぎ合わせた植物体。
【掲載論文】
タイトル:Defense-related callose synthase PMR4 promotes root hair callose deposition and adaptation to phosphate deficiency in Arabidopsis thaliana
著者:Kentaro Okada1,2, Koei Yachi1, Tan Anh Nhi Nguyen1, Satomi Kanno3, Shigetaka Yasuda1, Haruna Tadai1, Chika Tateda1, Tae-Hong Lee1, Uyen Nguyen4, Kanako Inoue1, Natsuki Tsuchida1, Taiga Ishihara1, Shunsuke Miyashima1, Kei Hiruma1, Kyoko Miwa5, Takaki Maekawa4, 6, Michitaka Notaguchi2,
Yusuke Saijo1
所属:1. 奈良先端科学技術大学院大学先端科学技術研究科 2.名古屋大学生物機能開発利用研究センター 3.名古屋大学高等研究院 4.ケルン大学植物科学研究所 5.北海道大学環境科学院 6.ケルン大学 CEPLAS Cluster of Excellence on Plant Sciences
掲載誌:The Plant Journal
DOI:10.1111/tpj.17134
【植物免疫学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses111.html
研究室ホームページ:https://bsw3.naist.jp/saijo/
(2024年11月19日掲載)
この記事をシェア ![]()