Research 研究成果の紹介
久保健一研究員、円谷徹之研究員、高山誠司教授らの研究グループが、近親交配を回避する受粉の新たな仕組みを解明
細胞間情報学講座の久保健一研究員、円谷徹之研究員、高山誠司教授らの研究グループが、植物が遺伝的に近い花粉を見分けて近親交配を回避する、全く新しい仕組みを発見しました。この研究成果は、米科学誌Science(11月5日号, Research Article)に掲載され、同号のPerspectivesに取り上げられました。また、日本経済新聞、産経新聞、日経産業新聞、日刊工業新聞、化学工業日報、奈良新聞、朝日新聞、読売新聞に記事として掲載されました。
プレスリリース詳細( 大学HP http://www.naist.jp/ 内コンテンツ )
高山誠司教授のコメント

記者会見の様子。写真奥から高山教授、久保研究員、円谷研究員自他を識別するには、「自己」を認識するか、「非自己」を認識するか、原理的には2つの方法が可能です。私達は、これまでアブラナ科植物が、自己の花粉の分子とだけ結合する雌しべの分子を使って、「自己」を認識していることを証明してきました。一方、今回の研究対象であるナス科・バラ科植物の花粉因子候補が見つかった際には、様々な状況証拠から「非自己」を認識しているというモデルを提唱しました。しかし、この正反対のモデルでは、数十種類は存在する「非自己」の雌しべの分子を、花粉の分子がどうやって認識できるのか、きちんと説明することが出来ませんでした。今回、花粉が、さらにその何倍もの認識分子を用意して「非自己」の雌しべの分子の認識にあたっていることを明らかにすることが出来ましたが、実際に認識し合う分子の組合せを見つけて、それらが相互作用することを実証するまで何年もかかりました。終始中心となって研究を進めてくれた久保健一研究員、円谷徹之研究員の執念とも言える粘りなしでは為し得なかった成果です。また、今回この成果を論文として発表できたのは、その他の共著者の方々、また論文にお名前を載せられなかった大勢の方々の御支援のおかげであります。この場を借りて深く感謝致します。
研究の概要
植物の多くは、虫や風などによって運ばれてくる様々な花粉の中から、自己や近縁の花粉を識別して排除する、自家不和合性と呼ばれる仕組みを発達させている。近親交配による悪い影響を回避するために、さらには種の遺伝的な多様性を保つために、植物にとって極めて重要な仕組みであると考えられている。
この自家不和合性の現象は、約250年程前から知られてきたが、植物がどの様にして「自己」と「非自己」を識別しているのかについては長年の謎であった。機構解明に向けた研究は1920年代の遺伝学的解析にスタートし、まず多くの植物の自家不和合性が、S遺伝子座上の複対立遺伝子(S1, S2, S3, ---, Sn)により制御されていることが示されてきた。すなわち、遺伝学的には、花粉と雌ずいが同じ番号のS複対立遺伝子を発現している時に不和合となると説明されてきた(図1)。
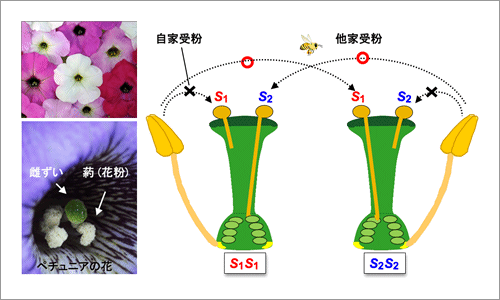
図1.実験材料のナス科植物ペチュニアの花(左)と自家不和合性の遺伝学的説明(右)
近年、様々な植物種を対象としてS遺伝子座の解析が進められ、まずアブラナ科植物において、多型性を持った花粉因子(SP11リガンド)と雌ずい因子(SRK受容体)の両者が、密接に連鎖する形でコードされていることが明らかにされた。さらに、同じS複対立遺伝子に由来するSP11とSRKが特異的に相互作用することにより、雌しべの細胞内で自家不和合性反応が誘起され、花粉の吸水・発芽が阻害されることが明らかになった。また、最近英国のグループにより、ケシ科植物のS遺伝子座には、アブラナ科植物とは全く異なる雌ずい因子(PrpSリガンド)と花粉因子(PrsS受容体)がコードされていることが明らかにされた。さらに、同一S複対立遺伝子に由来するPrpSが花粉表面のPrsSに特異的に結合すると、花粉が自殺(アポトーシス)することも示された。この様に、アブラナ科やケシ科植物では、使われる花粉因子と雌ずい因子の分子や機能は異なるものの、同じS複対立遺伝子上の両因子間の特異的な相互作用を介して「自己」を認識する点は共通であることが明らかにされてきた。
一方、ナス科やバラ科やオオバコ科の植物では、雌ずい因子としてS-RNaseというRNA分解酵素が利用されていることが20年程前から明らかにされ、これが花粉のRNAを分解する細胞毒として機能することが示唆されてきた。また、花粉因子の候補としてSLFというF-boxタンパク質が同定され、これがSCFユビキチンリガーゼの構成成分として機能する可能性が示唆されてきた。これら分子の性状から推定して、自家受粉時には花粉内に侵入してきたS-RNaseにより花粉管のRNAが分解されて花粉管の伸長が停止してしまうが、他家受粉の際には、花粉側因子SLFが侵入してきた非自己のS-RNaseを認識し、ポリユビキチン化し、分解することで無毒化している、というモデルが推定された。しかし、このモデルが正しいとすると、花粉因子SLFは、少なくとも数十種類は存在する自己と異なるS複対立遺伝子にコードされたS-RNaseをすべて認識する必要があり、果たしてその様な「非自己」認識が可能かどうか、疑問視されてきた。
今回、我々は、S複対立遺伝子上には、SLFに加え、SLFと類似した分子が少なくとも6種類コードされていることを明らかにした(Type-1〜6 SLFs)。さらに、この内の3種類のSLFs分子を用いて形質転換実験を行い、各SLF遺伝子を導入した花粉が、特定の非自己のS-RNaseを無毒化できる様に変化することを明らかにした。さらに、SLF分子と無毒化できるS-RNase分子とは、実際にタンパク質レベルで相互作用することを証明した。以上の結果をもとに、ナス科植物では、複数のSLF分子が協調的に機能することで、非自己の花柱に存在するすべてのS-RNase分子を無毒化できるという「協調的非自己認識」モデルを提唱した(図2)。
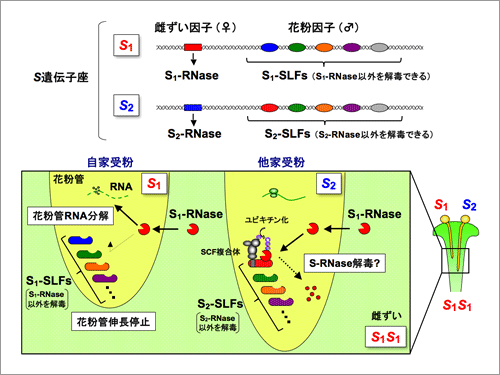
図2.ナス科植物の自家不和合性における「協調的非自己認識」モデル
ナス科植物の自家不和合性を制御するS遺伝子座には、雌ずい因子(S-RNase)と多数の花粉因子(SLFs)が密接に連鎖する形でコードされている(上)。雌ずいの通導組織に分泌されたS-RNaseは、伸長してくる花粉管に取り込まれ、花粉管のRNAを分解する細胞毒として機能し、自己花粉の伸長を停止させる(左)。花粉管で発現するSLFsは、いずれも「自己」の(同じS番号の)S-RNaseは認識しないが、「非自己」の(異なるS番号の)S-RNasesのいずれかを認識し、ユビキチン化し、無毒化(解毒)する。個々のSLFが認識しうる「非自己」のS-RNasesは限られるが、多数のSLFsが協調的に機能することで、すべての「非自己」S-RNasesの解毒、すなわち他家受精全般を可能とする(右)。
(2010年11月26日掲載)
この記事をシェア ![]()