Research 研究成果の紹介
マメ科植物の葉が動くしくみに迫る
葉の運動を駆動する細胞が伸展・収縮する際に細胞壁にあるスリットが開閉することを発見
~ダーウィンらが注目した植物の運動力の謎解明へ~
マメ科植物の葉が動くしくみに迫る
葉の運動を駆動する細胞が伸展・収縮する際に細胞壁にあるスリットが開閉することを発見
~ダーウィンらが注目した植物の運動力の謎解明へ~
【概要】
奈良先端科学技術大学院大学(学長:塩﨑 一裕)先端科学技術研究科 バイオサイエンス領域の中田 未友希助教、出村 拓教授らは、アカシア園芸の高原 正裕研究員、秋田県立大学の津川 暁助教、産業技術総合研究所の坂本 真吾主任研究員らと共同で、マメ科植物の葉の運動を駆動する細胞が、他の細胞にはない特殊な細胞壁構造を持つことを発見しました。
植物の運動は、1880 年に発表されたチャールズ・ダーウィンらの著作「The Power of Movement in Plants(植物の運動力)」においても取り上げられるなど、長い間多くの研究者を魅了してきました。そのなかでもマメ科は、昼に葉を開いて夜閉じる「就眠運動」をはじめとして、様々な葉の運動を示すことで有名な植物グループです。この運動は、葉の小葉や葉柄の付け根にある構造「葉枕(ようちん)」の細胞「運動細胞」が大きな伸展と収縮を繰り返すことで起こります。このような繰り返しの細胞伸縮は、硬い細胞壁に囲まれた植物細胞においては非常に稀な現象です。しかしながら、運動細胞がなぜ繰り返しの伸縮が可能なのかについては、これまでわかっていませんでした。
本研究では、繰り返しの伸縮を可能にするメカニズムに迫るため、運動細胞の細胞壁構造を、鮮明な高解像度の画像が得られる共焦点レーザー顕微鏡で詳細に調べました。その結果、運動細胞の細胞壁に切れ込み様の構造が多数存在することを発見しました。この切れ込み様の構造は、他の細胞の細胞壁にはない特殊な構造であったため、「葉枕スリット」と名付けました。葉枕スリットは、ダイズやインゲン、アズキのような作物から、おじぎ運動で有名なオジギソウ、クローバーやクズといった身近な雑草まで、葉の運動性を示すマメ科植物の葉枕に広く存在しました。詳細な細胞壁分析や、コンピューターシミュレーション、伸縮中の細胞における葉枕スリットの観察を組み合わせ、葉枕スリットが力学的に柔軟で、細胞の伸縮に伴って開閉する構造であることを明らかにしました。この研究の成果は、植物が葉の運動の進化の過程で、運動細胞を繰り返し伸縮させるために、細胞壁を特殊化することで切り紙のような伸縮しやすい構造を獲得した可能性を示す重要な発見です。
この成果は米国の植物生理学会の学会誌である Plant Physiology 誌オンラインサイトに 2023 年 2 月28 日(火)21 時に公開されます。(DOI:10.1093/plphys/kiad105)
【解説】
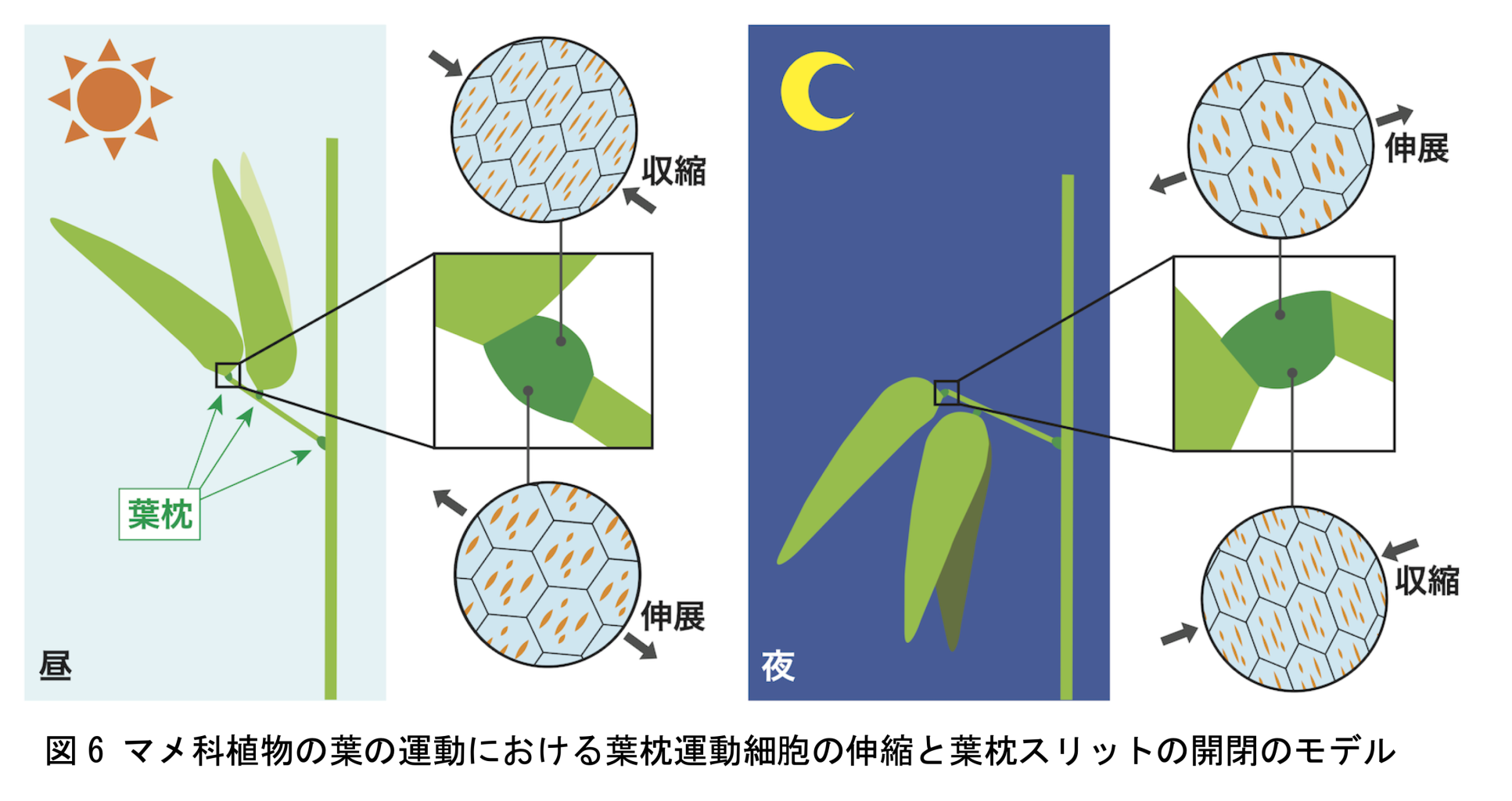
一部の植物グループで見られる葉が昼開き夜閉じる現象は、葉が夜に眠るように見えることから「就眠(しゅうみん)運動」と呼ばれています(図 1)。マメ科は、就眠運動をはじめとした、様々な葉の運動を示すことで有名な植物グループです。マメ科植物の葉が運動するとき、葉柄や小葉の付け根にあるとても小さい構造「葉枕(ようちん)」が変形して、蝶番(ちょうつがい)のような機能を果たします(図 2)。葉枕の上側と下側で、昼夜での伸縮が逆になることで葉の運動は駆動されています。例えば、上側が縮み、下側が伸びると、葉は起き上がります。反対に、上側が伸び、下側が縮むと、葉は垂れ下がります。葉枕の上下の伸縮において、葉枕の内部にある「運動細胞」と呼ばれる特殊な細胞群が、自身の細胞内圧を変化させることにより協調的に伸縮することがわかっています。運動細胞の伸縮は繰り返し、大きく、葉の長さ方向に偏って起こります(図 3)。このような繰り返しの細胞伸縮は、硬く変形しづらい細胞壁を持つ植物細胞においては非常に稀な現象です。本研究では、運動細胞がなぜ繰り返しの伸縮が可能なのか、どのように細胞伸縮の方向を制御しているのか、そのメカニズムに迫るため、葉枕の運動細胞の細胞壁に着目しました。
はじめに、大学構内に自生しているマメ科の雑草アレチヌスビトハギを採取し、共焦点レーザー顕微鏡による細胞壁の観察を行ないました。葉枕運動細胞の細胞壁セルロースを可視化したところ、セルロースをほとんど含まない、細胞伸縮の方向に対して垂直に走る、切れ込みのような構造が発見されました(図 4 左)。このような切れ込み様構造はこれまで他の植物細胞では観察された例がなく、葉枕の運動細胞に特有の細胞壁構造であると考えられたことから、「葉枕スリット」と名付けました。次に、葉枕スリットが、マメ科植物の葉枕で一般的に見られる構造かどうかを明らかにするため、様々なマメ科植物の葉枕の運動細胞で細胞壁の観察を行ないました。その結果、作物のダイズやインゲン、アズキ、おじぎ運動で有名なオジギソウ、身近な雑草であるクローバーやクズ、モデルマメ科植物であるミヤコグサなど、調べた計 13 種のマメ科植物全てで、同様の切れ込み様構造が観察されました。この結果から、葉枕スリットはマメ科植物が一般的に持つ、運動細胞に特有の細胞壁構造であると結論づけました。

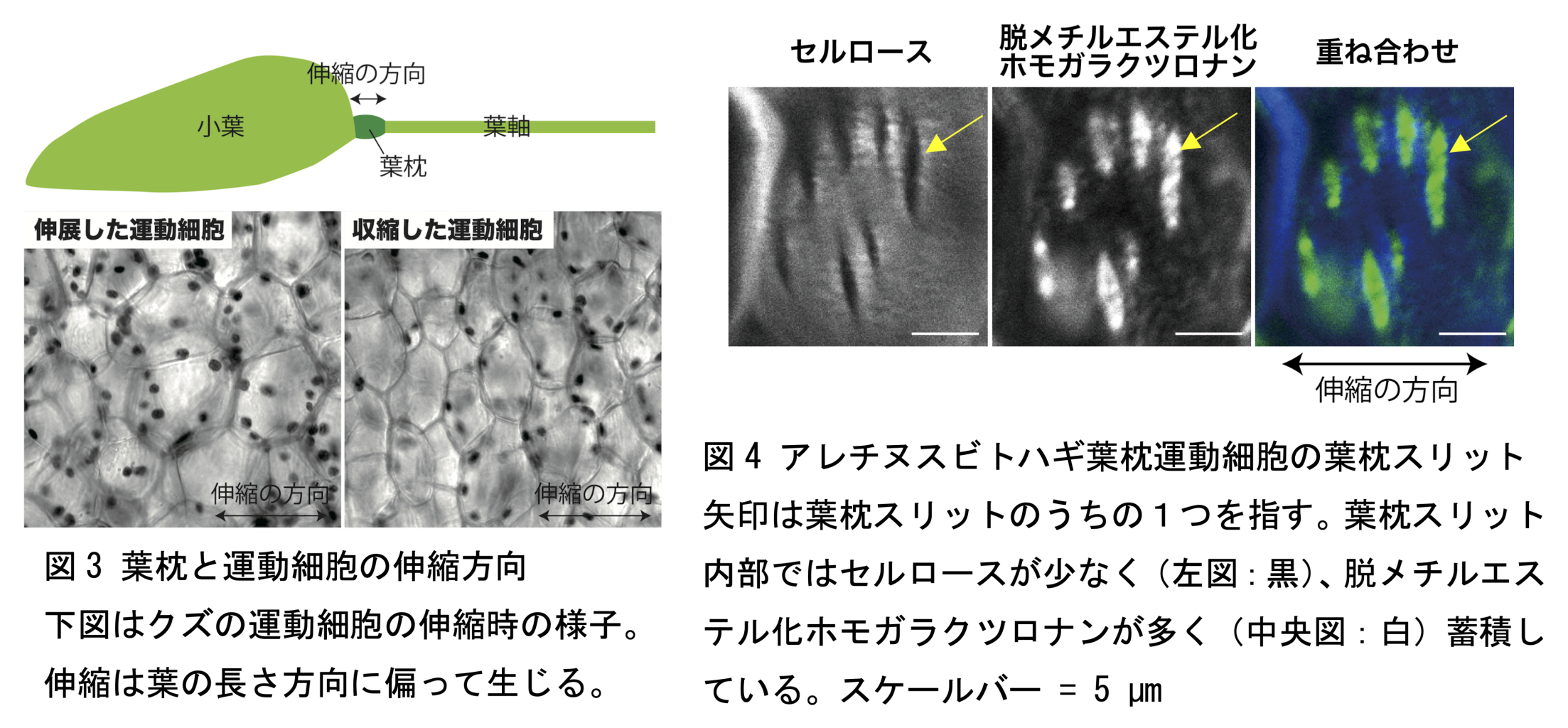
次に、葉枕スリットをさらに詳細に明らかにするため、セルロース以外の細胞壁多糖類の成分について、空間分布を、免疫染色により調べました。その結果、細胞壁の主成分であるペクチンの一種ホモガラクツロナンが、葉枕スリットと一致する空間分布を示すことがわかりました。特に、酵素によりメチルエステル基が除去された(脱メチルエステル化)状態のホモガラクツロナンは葉枕スリットの内部を充填するように蓄積していることがわかりました(図 4 中央、右)。この脱メチルエステル化ホモガラクツロナンは細胞壁を力学的に柔軟な構造にすることが知られています。細胞壁の構造から、葉枕スリットが力学的に柔軟な構造である可能性が考えられました。葉枕運動細胞における細胞壁の特殊化や、ホモガラクツロナンの重要性は、FTIR(Fourier Transform Infrared Spectroscopy: フーリエ変換赤外分光法)を用いた細胞壁分析や、UPLC(Ultra-Performance Liquid Chromatography: 超高パフォーマンス液体クロマトグラフィー)を用いた単糖分析によって裏付けられました。さらに、コンピュータシミュレーションの一種である FEM(Finite Element Method:有限要素法)により、細胞壁に力学的に柔軟なスリットが多数、同じ方向に並んで存在することが、細胞の伸展方向の制御に重要であると推定されました。
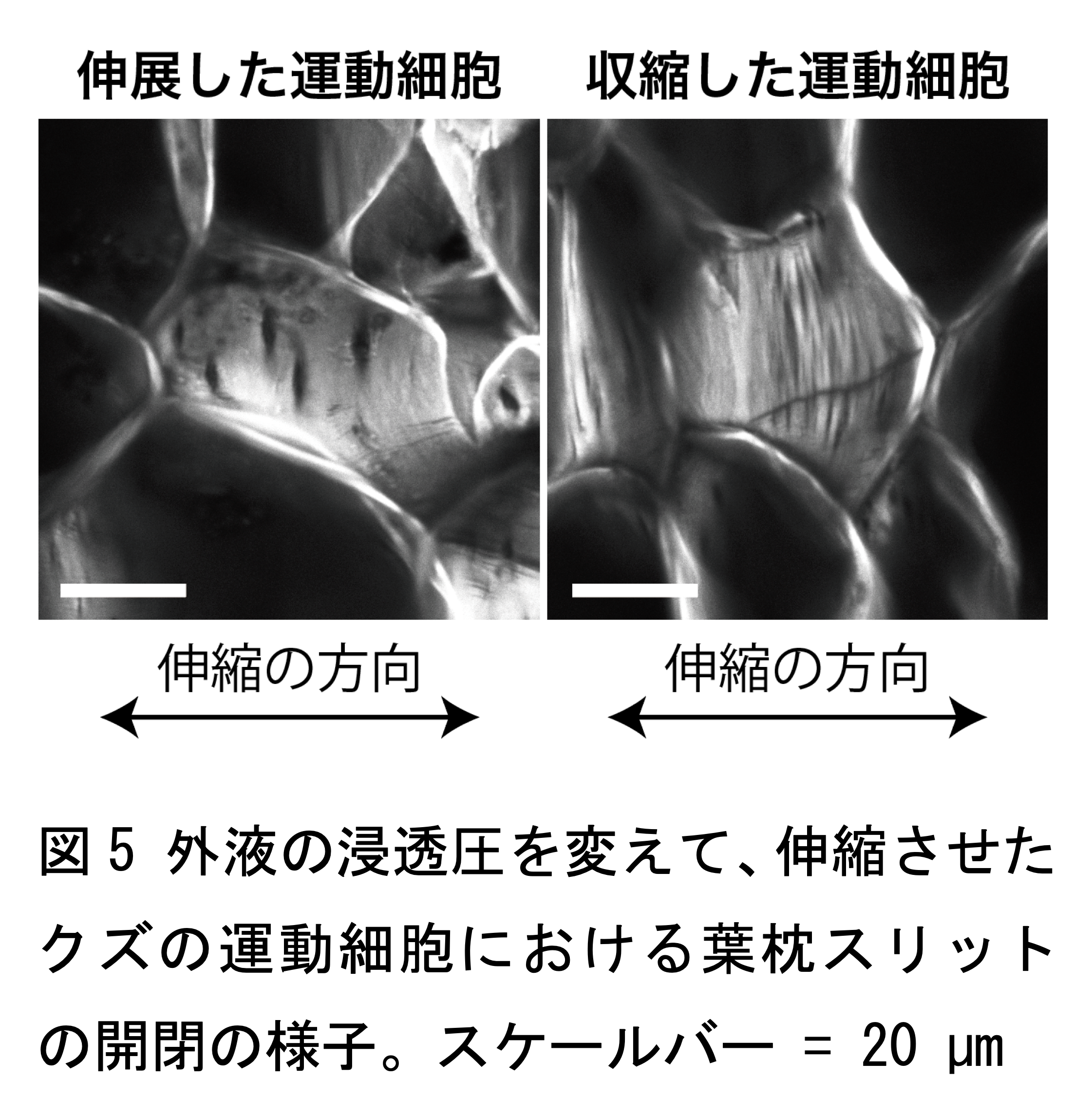
最後に、葉枕スリットが力学的に柔軟な構造なのかを実験的に検証するため、マメ科植物のクズを材料とし、葉枕組織片を切り出して、外液の浸透圧を変えることにより強制的に細胞を伸縮させる実験を行いました。その結果、伸展した細胞では葉枕スリットが開いており、収縮した細胞では閉じていることがわかりました(図 5)。また、同一細胞の伸縮前後の細胞の比較から、葉枕スリットは細胞の伸縮中に裂けることなく開閉する構造であることがわかりました。
これらの研究結果から、マメ科植物の葉枕運動細胞には、特有の柔軟な細胞壁構造「葉枕スリット」が存在すること、運動細胞の伸縮方向の制御に関わる可能性があること、運動細胞の伸縮に伴って葉枕スリットが開閉することが明らかとなりました。また、運動細胞の伸縮により葉の運動が駆動されることから、葉枕スリットは葉の運動中にも開閉していると推定されます(図 6)。
【背景と目的】
マメ科植物の葉の運動を含め様々な植物の運動が、1880 年に発表されたチャールズ・ダーウィンらの著作「The Power of Movement in Plants」において記述されるなど、長い間多くの研究者を魅了してきました。茎の回旋運動や、光や重力に反応して茎や胚軸を曲げる屈性など、植物は成長に伴って様々な運動性を見せます。一方、マメ科植物の葉の運動は、成長を伴わない点が、他の運動とは根本的に異なっています。マメ科植物では、葉柄や小葉の付け根にある葉枕に存在する運動細胞が、繰り返し大きく伸縮することで葉の運動を駆動しています。植物細胞は一般的に硬い細胞壁で覆われており、成長に伴って細胞壁を変質させて大きく伸びることはあっても、大きく縮むことは稀です。運動細胞が他の細胞と大きく違う点は、その可逆的な伸縮能です。このことから、運動細胞の細胞壁は、一般的な細胞とは異なる性質を持つのではないかと推察されますが、運動細胞と他の細胞の細胞壁の違いについては多くが未解明でした。そこで、本研究では運動細胞の細胞壁を詳細に明らかにすることを目的としました。
【今後の展開】
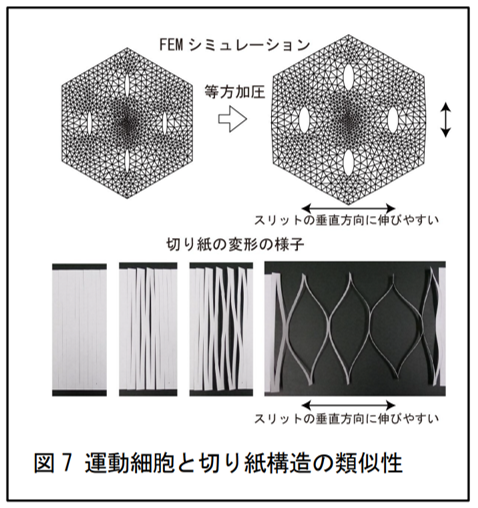
本研究の成果から、運動細胞の繰り返しの伸縮は、細胞壁の中に多数の柔軟なスリット構造が同じ方向に並んで存在することにより可能となっていることが推察されました。興味深いことに、運動細胞における葉枕スリットの役割は、局所的な破壊を伴うことなくシート状構造の一方向への伸展性を高めるという点で、切り紙構造におけるスリットの役割と類似しています(図 7)。また、進化的な観点からは、植物が葉の運動性を獲得する進化の過程で、細胞壁の構造や機能を特殊化してきた様子がうかがえます。マメ科は、現存する被子植物の中で特に繁栄していると考えられている植物グループの 1 つです。この点を踏まえると、今後細胞壁の特殊化による葉の運動性獲得過程が明らかになることにより、植物の生存戦略の進化と多様性のさらなる理解につながることが期待されます。また将来的に、細胞壁の特殊化の分子メカニズムが明らかとなった場合、ゲノム編集技術などと組み合わせることで、植物の持つ細胞壁形成制御システムを活用したバイオ新素材の開発などに貢献する可能性もあります。
【用語解説】
マメ科植物:780 属約 2 万種で構成される、被子植物の中で 3 番目に大きな科である。マメ科には様々な有用植物が属しており、ダイズやインゲン、アズキ、エンドウ、ラッカセイなどの作物をはじめ、ローズウッドやファルカタ、アカシアといった材木として利用される樹木、スイートピーやフジ、ハギ、ルピナス、エバーフレッシュ、オジギソウといった園芸植物、カンゾウやエビスグサなどの薬草が含まれる。また、クローバー(シロツメクサ)やレンゲソウ、クズ、ネムノキなど身近な植物も多い。
葉枕(ようちん):マメ科植物など一部の植物グループが持つ、葉の葉柄や小葉の付け根にある膨らんだ構造。
共焦点レーザー顕微鏡:レーザー光照射と光学系の工夫により、比較的分厚い試料からボケの少ない光学切片像として蛍光を高解像で(細胞レベルで)観察することを可能とする顕微鏡。
細胞壁:植物の細胞を取り囲む、セルロースなどの多糖を主成分とする硬い構造。細胞の伸縮の程度や方向を制御している。食料と競合しないバイオ資源として、産業的にも重要である。
脱メチルエステル化ホモガラクツロナン:ホモガラクツロナンは細胞壁を構成する多糖であるペクチンの一種。カルシウム架橋されていない脱メチルエステル化ホモガラクツロナンは、力学的に柔軟であることが知られている。
FTIR(フーリエ変換赤外分光法):赤外線の吸収スペクトルから、化合物の構造に関して定量・定性分析を行う装置。
UPLC(超高パフォーマンス液体クロマトグラフィー):液体クロマトグラフィーの一種。溶液中の物質を、物性の違いを利用して分離・定量する装置。
FEM(有限要素法):主に時空間的な変動を予測する数値解析手法。構造解析シミュレーションでは弾性体を有限要素に分けて物体にかかる力・変位などを計算することができる。
本研究は、文部科学省の科学研究費補助金 (JP20K15832、JP19K16174、JP19K23753、JP20K06707、JP18H05484、JP18H05489)と科学技術研究機構(CREST(JPMJCR2121))の支援を受けて行われました。
【論文情報】
タイトル:Pulvinar slits: Cellulose-deficient and de-methyl-esterified pectin-rich structures in a legume motor cell
著 者 名 :Masahiro Takahara, Satoru Tsugawa, Shingo Sakamoto, Taku Demura, Miyuki T Nakata
雑誌:Plant Physiology
DOI:10.1093/plphys/kiad105
【植物代謝制御研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses104.html
研究室ホームページ:https://bsw3.naist.jp/demura/
中田未友希助教:http://www.naist.jp/gender/contents/message/interview/28.html
(2023年03月07日掲載)
この記事をシェア ![]()