Research 研究成果の紹介
花を咲かせないようにする仕組みを発見
花成ホルモン「フロリゲン」と抑制因子が共通のパートナーを奪い合い
開花時期の操作や食糧増産に期待
花を咲かせないようにする仕組みを発見
花成ホルモン「フロリゲン」と抑制因子が共通のパートナーを奪い合い
開花時期の操作や食糧増産に期待
【概要】
奈良先端科学技術大学院大学 (学長: 横矢直和) 先端科学技術研究科 バイオサイエンス領域 花発生分子遺伝学研究室の山口暢俊助教と米国ペンシルバニア大学 Doris Wagner(ドリス・ワグナー)教授らの国際共同研究グループは、植物が葉をつくる成長をしているときに、花をつくる遺伝子の働きを OFF にしておく詳細な仕組みを世界に先駆けて明らかにしました。この成果により、開花の時期を人為的に調節できるようになれば、食糧の増産や安定供給などが期待できます。
花を咲かせるためには、花芽の形成を開始する小さなタンパク質「フロリゲン」が葉で作られた後、茎の先端(茎頂)へと運ばれて働くことがわかっていました。このフロリゲンは茎頂に移動後、遺伝子発現のスイッチを ON にする特定のパートナーの因子(転写因子)と結合することにより、花をつくる遺伝子に働きかけます。しかし、日の長さや気温など花を咲かせるために適した環境条件でない場合には、まだ花を咲かせずに待ったほうが生存や適応にとって有利になります。そのためには、フロリゲンが働かないようにブロックする必要があります。これまでに、フロリゲンの働きを抑制するタンパク質が見つかっていましたが、「その抑制因子がどのようにフロリゲンの作用をブロックするのか?」という詳しい仕組みについては謎でした。
山口助教らの国際共同研究グループは、モデル植物のシロイヌナズナを使って実験を重ねた結果、フロリゲンは、その抑制因子と対抗し、パートナーの転写因子を奪い合うことを突き止めました。このことから、フロリゲンがパートナーの因子を奪われてしまうと花をつくる遺伝子に働きかけることができなくなることを明らかにしました。花の形づくりを進めるかどうかを決める仕組みを解明することは、植物の進化や生き残り戦略を知るうえでも非常に重要です。本研究の成果は 2020 年 10 月 12 日付けで Nature Communications(オンラインジャーナル)に掲載されました。
山口暢俊助教のコメント

アメリカに留学していた時、私は花の運命を決めるLEAFY(LFY)遺伝子の研究を行っていました。植物は日の長さなど外的な環境を読み取って、LFY遺伝子を発現させて、花の運命を決定します。私が行った研究により、LFYが働く仕組みはよくわかっていました(Yamaguchi et al., 2013 Dev Cell; Yamaguchi et al., 2014 Science; Wu and Yamaguchi e-life)。ですが、LFY遺伝子を働かないように抑える仕組みは不明でした。私は花成ホルモン「フロリゲン」と逆の働きを持つTERMINAL FLOWER1(TFL1)に注目し、LFY遺伝子の働きを直接的に抑制することを見出していました。その後、ドリスワグナー博士の指導のもと、Yangくんが中心になって、この研究を完成させてくれました。フロリゲンとTFL1による花を咲かせる仕組みの解明は学術的に重要なだけではなく、農学や園芸という分野においても大きな影響を与えるものです。
【背景】
植物は環境からの様々な情報を利用して花を咲かせる適切なタイミングを決めています。特に光に
応答したタイミングの決定において重要な役割を果たしているのが、花成ホルモン「フロリゲン」と呼ばれる FLOWERLING LOCUS T (FT)タンパク質です。花を咲かせるように仕向ける光の環境条件で育てた植物は、まず葉でフロリゲンを産出します。そのフロリゲンは篩管を通って茎頂へと運ばれ、パートナーである遺伝子発現のスイッチを ON にする転写因子 FD と結合し、花芽をつくる遺伝子に働きかけます。
しかし、光など環境の情報が花を咲かせるのに適した条件でない場合には、茎頂での花芽形成は抑制されています。この抑制を行っているのが、TERMINAL FLOWER 1(TFL1)タンパク質です。このTFL1タンパク質も FT のパートナーである FD に結合して複合体を形成することがわかっていましたが、実際にその TFL1-FD タンパク質複合体がどのような役割を持つ遺伝子の発現を変化させて、どのようにFT の働きを抑えるのか?という詳しい仕組みは未解明のままでした。
【研究手法と成果】
はじめに私たちは、花を咲かせるのを抑える TFL1 と FD の複合体に注目しました。花を咲かせるのに適していない条件で、TFL1 とそのパートナーの FD が直接結合する遺伝子を「クロマチン免疫沈降法」という免疫の反応を使う手法によって解析しました。クロマチン免疫沈降法では、タンパク質とDNA の相互作用を網羅的に同定することができます。その結果、TFL1 と FD の複合体は 2372の共通の遺伝子に直接結合することがわかりました(図 1(a))。それらの遺伝子の中には、光に応答して花を咲かせるように働く遺伝子( GIGANTIEA ・ CONSTANS ・ SUPPRESSOR OF CONSTANS1 遺伝子)や細胞に花の性質を付与する遺伝子(LEAFY(LFY)・APETALA1・FRUITFULL遺伝子)が含まれていました(図 1(b))。さらに、FD の機能がなくなった場合、TFL1の遺伝子に対する結合もなくなることから、TFL1は FD をパートナーとして遺伝子に働きかけることを明らかにしました。
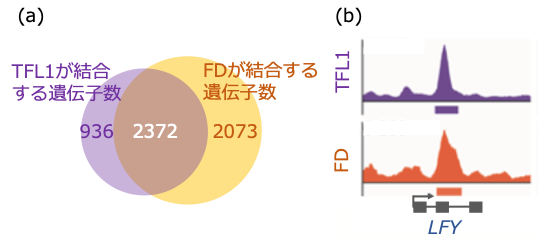
図 1.TFL1 と FD 複合体より制御される遺伝子の同定
(a) TFL1 と FD の結合部位の網羅的同定
(b) LFY 遺伝子に対する TFL1 と FD の結合
TFL1 と FD の複合体が遺伝子の働きを抑制する詳細な仕組みを明らかにするために、LFY 遺伝子の発現制御に注目しました。これまでに TFL1 タンパク質が働かなくなった突然変異体では、LFY 遺伝子が発現する花芽だけでなく、本来発現しない茎頂でも発現することがすでに報告されていました。そのため、TFL1 は LFY 遺伝子を抑制するのに必要であると考えられていましたが、TFL1 は転写抑制をする因子を積極的に呼び込んで転写抑制を行うのか、それとも受動的に発現を抑制するのかはわかっていませんでした。そこで、TFL1 タンパク質をステロイドホルモンの作用により、植物細胞の核に移行させる実験を行ったところ、すばやく LFY 遺伝子の転写が抑制されました。このことから、TFL1 とFD の複合体は、積極的に転写抑制を行うことがわかりました。
さらに、クロマチン免疫沈降法により、TFL1 と FD の複合体は LFY 遺伝子の塩基配列の中で 2番目のエキソン(遺伝情報として意味がある配列)部分に結合することがわかりました。そこでまず、LFY遺伝子の第 2 エキソンに TFL1 と FD の複合体が結合する配列が存在するかどうかを調べたところ、LFY 遺伝子の第 2 エキソンには、3 つの FD が結合する配列があり、それらの配列はシロイヌナズナだけでなく、近縁の植物でも高度に保存されていました(図 2(a))。これらの 3 つの配列に変異を導入すると、TFL1 がそこには結合しなくなることがわかりました。その影響により、LFY 遺伝子が本来発現する花芽ではなく、茎頂で異所的に発現するようになりました(図 2(b))。これらの結果から、TFL1と FD の複合体は、LFY 遺伝子の第 2 エキソン上に存在する結合配列に結合し、その転写を抑制することがわかりました。
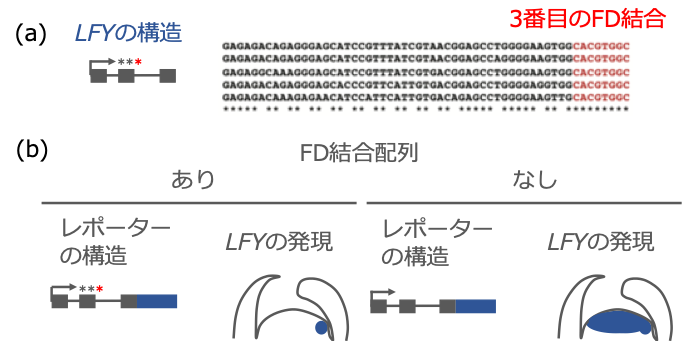
図2. LFY 遺伝子の第 2 エキソンにある TFL1 と FD の結合配列の役割
(a) LFY 遺伝子の第 2 エキソンにある結合配列の保存性
(b) 結合配列に変異を導入したレポーターと LFY 遺伝子の発現
TFL1 だけでなくフロリゲンの分子実体である FT タンパク質も、FD をパートナーとしてDNA に働きかけます。そこで、FT も LFY 遺伝子を制御するかどうかを調べたところ、花を咲かせる直前にFT の発現を人為的に抑制した場合、第 2 エキソン上に存在する結合配列の機能に依存して LFY 遺伝子の発現は下がることがわかりました。さらに、この発現の変化によって花の性質が弱くなり、花が咲くのが遅くなることも明らかにしました。TFL1 と FT が逆の制御をする遺伝子が LFYであると判明したため、両者がこの遺伝子座上で拮抗するメカニズムを調べました。FT を薬剤や光によって発現誘導させると、TFL1 遺伝子の発現量や FD の LFY 遺伝子座における結合は変わらず、一方でTFL1の LFY 遺伝子座における結合のみが減ることを突き止めました。そのため、TFL1 と FT の拮抗作用を支える仕組みが、パートナーFD の競合であることを明確にすることに成功しました (図 3)。
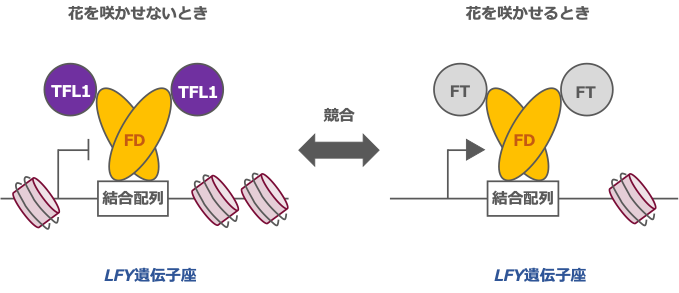
図 3. 花を咲かせないときと咲かせるときの仕組みの違い
花を咲かせないときには、TFL1-FD 複合体が花をつくる遺伝子の働きを抑えている。TFL1
と FT は競合し、FT-FD 複合体が形成されると花をつくる遺伝子が働き、花の形づくりが実
行されていく。
パートナーFD の競合による TFL1 と FT の拮抗作用により、植物が花を咲かせずに待つのか?それとも花を咲かせるかを決定します。そのあと、LFY 遺伝子が細胞に花の性質を付与して、花の形づくりが実行されていきます。
【波及効果】
フロリゲンの分子的な実体やそのホルモンの受容体の役割、およびフロリゲンが働く細胞の可視化など、フロリゲンが働いて花づくりが進んでいく仕組みはよくわかっていました。今回、そのフロリゲンによる花づくりに「待ったをかける」分子的な仕組みを世界にさきがけて発見しました。また、花を咲かせる仕組みと咲かせない仕組みを知ることで、最適に人為調節できる可能性があり、食糧増産や安定供給が期待されます。
【研究プロジェクトについて】
本成果は日本学術振興会(JSPS)科学研究費補助金、学振海外特別研究員支援を受けて行いました。
【掲載論文】
タイトル: TERMINAL FLOWER 1-FD complex target genes and competition with FLOWERING LOCUS T
著者: Yang Zhua, Samantha Klasfelda, Cheol Woong Jeonga, Run Jina, Koji Gotob, Nobutoshi Yamaguchia,c, Doris Wagnera.
所属: a. ペンシルバニア大学、 b.岡山県農林水産総合センター、 c.奈良先端科学技術大学院大学
掲載誌: Nature Communications
DOI: 10.1038/s41467-020-18782-1
【用語解説】
茎頂: 茎の先端部にあり、葉や花を作る組織。
フロリゲン: 植物が日長の変化に応じて花を咲かせる時に、葉で合成され、茎頂へと運ばれ花の形成を促す物質。シロイヌナズナでは、FT タンパク質が実体であることがわかっている。
転写因子: 遺伝子の発現の ON、OFF を切り替えるタンパク質。DNA 上の特定の結合配列に結合し、遺伝子の発現量を調節する。
DNA: 細胞の核内で長い鎖状の 2 重らせん構造を形成し、生物の遺伝情報を保持している物質。
シロイヌナズナ: 遺伝子の解析を行うのに適したアブラナ科の 1 年草。
FD タンパク質: 茎頂で発現するジッパーのような分子構造(bZIP 型)の転写因子。葉から運ばれてきた FT タンパク質と複合体を形成する。
TFL1 タンパク質: ホスファチジルエタノールアミン結合タンパク質ファミリーに属する球状のタンパク質。FT タンパク質も同じファミリーに属するが、FT とは異なり花を咲かせないように働く。
クロマチン免疫沈降法:抗体を用いて、タンパク質と DNA との結合を検出する方法。
LFY 遺伝子: 植物特有の転写因子をコードする遺伝子。細胞に花の性質を付与する働きがある。
エキソン: 遺伝情報がコードされている翻訳配列。遺伝情報がコードされていない配列はイントロンと言う。
【原著論文URL】
https://www.nature.com/articles/s41467-020-18782-1
【花発生分子遺伝学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:https://bsw3.naist.jp/ito/
(2020年10月15日掲載)
この記事をシェア ![]()