Research 研究成果の紹介
バイオサイエンス研究科分子免疫制御研究室の川﨑拓実助教、河合太郎准教授らが、ウイルス感染に対抗する新たな免疫反応の仕組みを解明
バイオサイエンス研究科分子免疫制御研究室の川﨑拓実助教、河合太郎准教授らは、ウイルス感染に対する免疫反応が、細胞膜に存在するリン脂質の一種ホスファチジルイノシトール5リン酸[1]により制御されていることを明らかにした。
河合太郎准教授のコメント
 今回の成果は、ウイルス感染に伴い生じるI型インターフェロンの産生が、細胞内脂質の一種ホスファチジルイノシトール5リン酸により制御されていることを明らかにしたものです。興味深いことに、合成したホスファチジルイノシトール5リン酸をマウスの生体に投与すると、抗体と呼ばれる免疫物質の産生を誘導することが分かりました。このことから、ホスファチジルイノシトール5リン酸は、インフルエンザウイルス等に対する免疫賦活剤の開発に繋がることが期待されます。本研究は、本研究室の川﨑拓実助教が中心となり行ったものです。また、大阪大学審良静男先生、同Daron Standley先生との共同研究であり、最先端研究開発支援プログラムの支援を受け行われました。この場を借りて感謝申し上げます。
今回の成果は、ウイルス感染に伴い生じるI型インターフェロンの産生が、細胞内脂質の一種ホスファチジルイノシトール5リン酸により制御されていることを明らかにしたものです。興味深いことに、合成したホスファチジルイノシトール5リン酸をマウスの生体に投与すると、抗体と呼ばれる免疫物質の産生を誘導することが分かりました。このことから、ホスファチジルイノシトール5リン酸は、インフルエンザウイルス等に対する免疫賦活剤の開発に繋がることが期待されます。本研究は、本研究室の川﨑拓実助教が中心となり行ったものです。また、大阪大学審良静男先生、同Daron Standley先生との共同研究であり、最先端研究開発支援プログラムの支援を受け行われました。この場を借りて感謝申し上げます。
研究の概要
【研究の背景】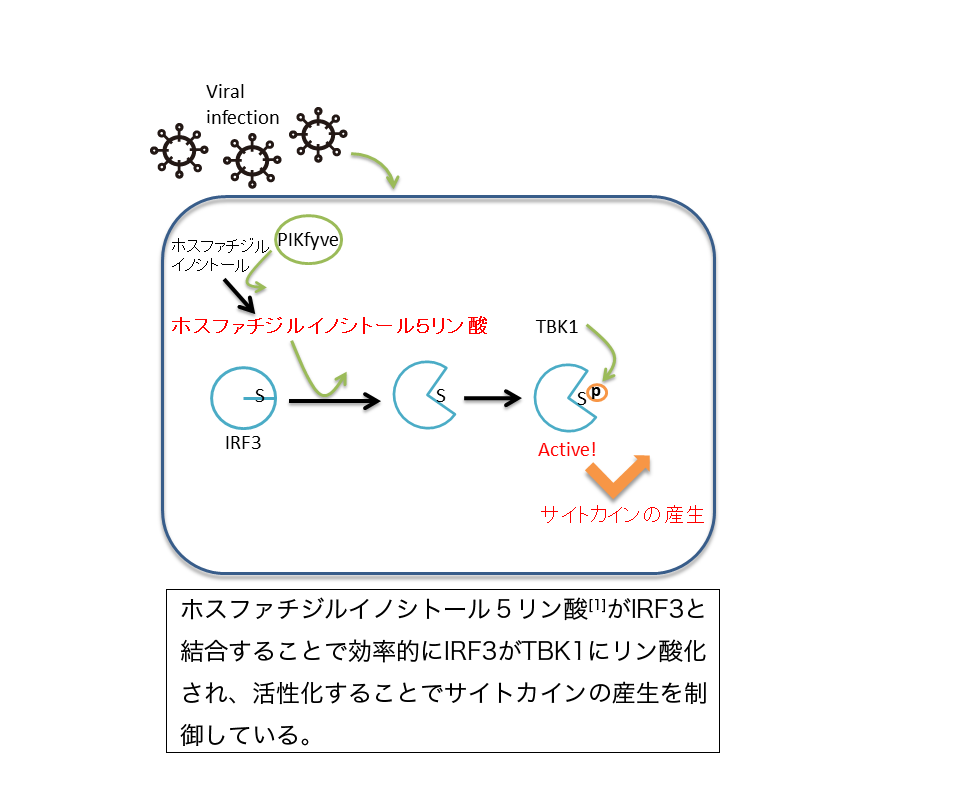
ウイルスが細胞に感染すると、細胞はI型インターフェロンや炎症性サイトカインといった液性因子(サイトカイン)を産生することでウイルスを排除しようとします。これらのサイトカインは引き続きT細胞やB細胞などにより担当される獲得性免疫の誘導に重要な役割を果たします。これまで、我々のグループでは、自然免疫とよばれる、病原体の認識からサイトカイン産生に至る過程の研究を進めていました。特に、ウイルス感染の認識からインターフェロン産生に至る細胞内のシグナル伝達において、リン酸化酵素TBK1[2] (Tank binding kinsae1)よる転写因子[3]IRF3のリン酸化が必須であることを明らかにしてきました。しかし、このTBK1-IRF3シグナルがどのように活性化するのか不明でした。
【本研究成果が社会に与える影響(本研究成果の意義)】
川﨑助教、河合准教授、審良教授らのグループは、TBK1-IRF3活性化のスクリーニング系を立ち上げ、その結果、ホスファチジルイノシトール5リン酸[1]がTBK1-IRF3シグナルを増強することを突き止めました。細胞にウィルスをはじめとする病原体が感染すると細胞内のホスファチジルイノシトール5リン酸レベルが増加することを見出しました。ホスファチジルイノシトール5リン酸はIRF3が結合し、さらにこの結合によりIRF3はTBK1により効率よくリン酸化されることがわかりました。その結果IRF3は活性化し、インターフェロンを含むサイトカインの産生を誘導することがわかりました。また、合成したホスファチジルイノシトール5リン酸を抗原と共にマウスに投与することで、抗原特異的な抗体産生を誘導したことから、今後、ホスファチジルイノシトール5リン酸[1]を用いた効果的なワクチン開発につながることが期待できます。
【特記事項】
本成果は、雑誌「Cell Host Microbe」オンライン版に掲載されました。なお本研究は、内閣府/日本学術振興会・最先端研究開発支援プログラムの支援を受けて行われました。
【解説】
<研究の背景>
ウイルスなどの病原体の感染は自然免疫系により認識されることが知られています。この認識により、細胞はインターフェロンや炎症性サイトカインをはじめとする液性因子(サイトカイン)を産生します。細胞から放出されたサイトカインは引き続きキラーT細胞の活性化やB細胞による抗体産生といった獲得性免疫による生体防御の誘導に重要な役割を果たします。自然免疫による病原体の認識は、病原体固有に存在する構造を認識する受容体(自然免疫受容体)により行われます。自然免疫受容体が病原体成分を認識すると細胞内シグナルシグナル伝達たんぱく質が順次活性化し、最終的に転写因子が活性化します。その結果、サイトカイン遺伝子の発現が誘導されて、サイトカインの産生が開始されます。各々の病原体由来の構造物に対する自然免疫受容体と、受容体からサイトカイン産生に至るシグナル伝達経路の全容は明らかになりつつありますが、依然として不明な点も数多く残されています。TANK Binding kinase 1(TBK1)は、ウイルス感染により活性化し、転写因子Interferon Regulatory Factor(IRF)3 を直接リン酸化するリン酸化酵素として知られています。リン酸化されたIRF3は細胞質内より核内に移行し、I型インターフェロンをはじめとするサイトカインの発現を誘導します。私たちはTBK1によるIRF3のリン酸化がウイルスに対するI型インターフェロン産生に必須であることを明らかにしてきましたが、どのようにTBK1-IRF3シグナルの活性化が制御されているかは不明でした。そこで私たちは、TBK1-IRF3シグナル活性化メカニズムを明らかにするために研究を行いました(図1)。
<研究手法と成果>
私たちはTBK1-IRF3の活性化メカニズムを明らかにするため、TBK1-IRF3のシグナルを活性化し得る細胞内因子の探索を行いました。その結果、ホスファチジルイノシトール5リン酸がIRF3と結合すること、この結合はTBK1によるIRF3のリン酸化を増強することで、インターフェロン産生を促進することを突き止めました。また、イノシトールリン酸の5位をリン酸化することでホスファチジルイノシトール5リン酸の産生に関わるリン酸化酵素PIKfyveの活性化を薬剤により阻害したり、ノックダウン法により発現を抑制すると、ウイルス感染によるインターフェロンの産生が減少することがわかりました。ウイルス感染によりホスファチジルイノシトール5リン酸の細胞内の量は増えることも分かりました。また、このホスファチジルイノシトール5リン酸の産生はPIKfyveの阻害により抑制されることがわかりました。以上のことから、ウイルス感染時の細胞の応答には、ホスファチジルイノシトール5リン酸が必要であることがわかりました。さらに、樹状細胞に合成ホスファチジルイノシトール5リン酸を与えると、インターフェロンなどのサイトカインが産生されることがわかりました。ホスファチジルイノシトール5リン酸が樹状細胞を活性化することができることから、ホスファチジルイノシトール5リン酸が生体内で抗体を作る時の補助的な役割「免疫賦活化能」を発揮するか調べました。ホスファチジルイノシトール5リン酸を抗原とともにマウスに免疫した結果、抗原特異的な抗体が産生することから、ホスファチジルイノシトール5リン酸に免疫賦活化能があることがわかりました(図2)。
<今後の期待>
ホスファチジルイノシトール5リン酸は、ホスファチジルイノシトール3リン酸や、ホスファチジルイノシトール4リン酸などに比べ、役割があまり知られていないイノシトールリン脂質であり、生体内での存在量も少ないのです。今回はじめて、自然免疫のシグナル伝達において、ホスファチジルイノシトール5リン酸がウイルスに対する免疫応答で重要な役割をもつことが明らかになりました。また、ホスファチジルイノシトール5リン酸が免疫賦活化能を有していることから、新しいタイプの免疫賦活剤として活用できる可能性があります。ホスファチジルイノシトール5リン酸は、これまでの免疫賦活剤と異なり、生体内由来の物質であり、より毒性が少なく安全性の高い免疫賦活化剤としての活用が期待されます(図3)。
<補足説明>
[1] イノシトールリン脂質、ホスファチジルイノシトール5リン酸
イノシトールリン脂質とはイノシトール環と脂肪酸からなるリン脂質。イノシトールリン脂質の内、イノシトール環の5位の部位がリン酸化されたものをホスファチジルイノシトール5リン酸と呼ぶ。
[2] リン酸化酵素
ATPからリン酸基を基質分子に転移する酵素の総称であり、基質はリン酸基の転移により活性化の調節を受ける。
[3] 転写因子
遺伝子の働きをオンにしたりオフにしたりする機能を持つタンパク質。DNA上に存在する転写を制御する領域に結合し、遺伝子発現のタイミングや量を調節する。
[4] 樹状細胞
抗原提示細胞として機能する免疫細胞。また、積極的にサイトカイン等を放出することにより、生体防御に役立っている。
<著者・掲載論文・雑誌>
- Takumi Kawasaki, Naoki Takemura, Daron M. Standley, Shizuo Akira and Taro Kawai
- The second messenger phosphatidylinositol-5-phosphate facilitates antiviral innate immune signaling
- Cell Host Microbe
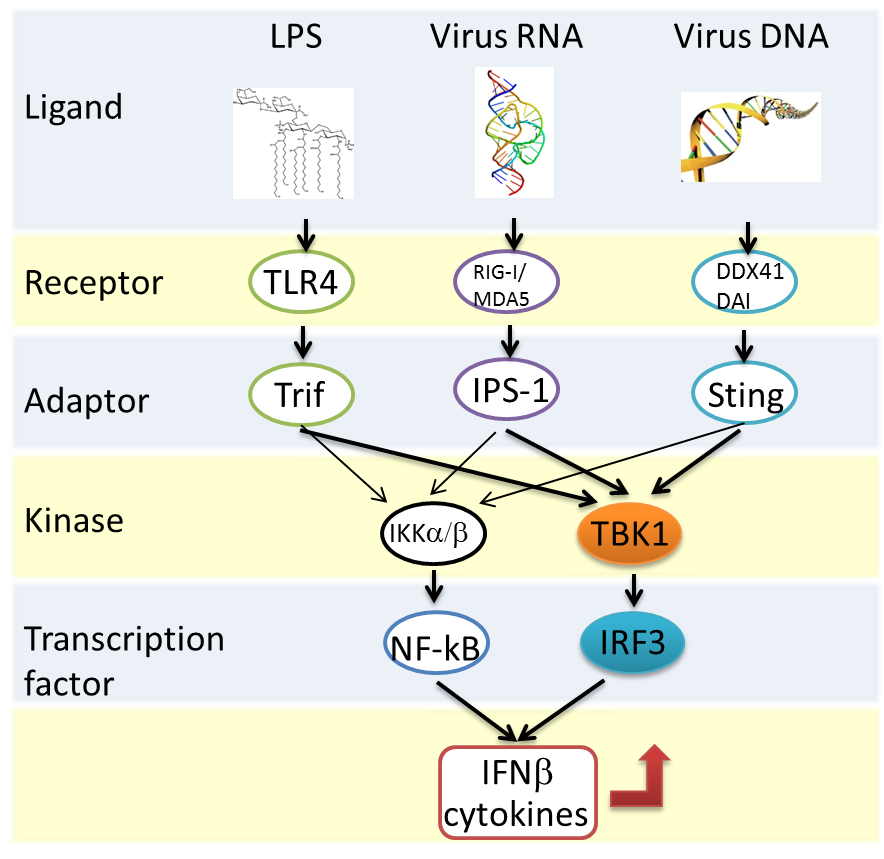
図1 自然免疫のシグナル伝達
ウイルスなどの病原体由来の構造物が、細胞内のレセプターに認識されると、各々のアダプターたんぱく質を介して、TBK1-IRF3シグナルなどが活性化し、インターフェロンなどのサイトカインの産生を誘導する。
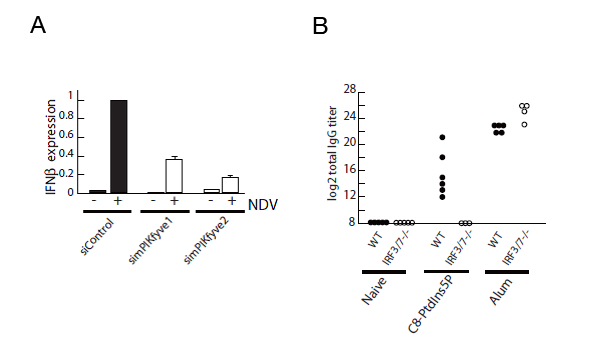
図2 ホスファチジルイノシトール5リン酸が自然免疫の活性化に重要である
A, ホスファチジルイノシトール5リン酸の産生に関わるリン酸化酵素PIKfyveの発現を抑制するとウイルス(Newcastle disease virus; NDV)感染によるインターフェロン(IFNβ)が減少する。
B, マウスに合成ホスファチジルイノシトール5リン酸(C8-PtdIns5P)もしくはAlum(陽性コントロール)とともに卵白アルブミン(OVA)抗原を投与して、OVA特異的抗体(IgG)の量を計ると、野生型マウス(WT; Wild type)ではホスファチジルイノシトール5リン酸により抗体産生が上昇するものの、IRF3/7ノックアウトマウスでは抗体産生しない。
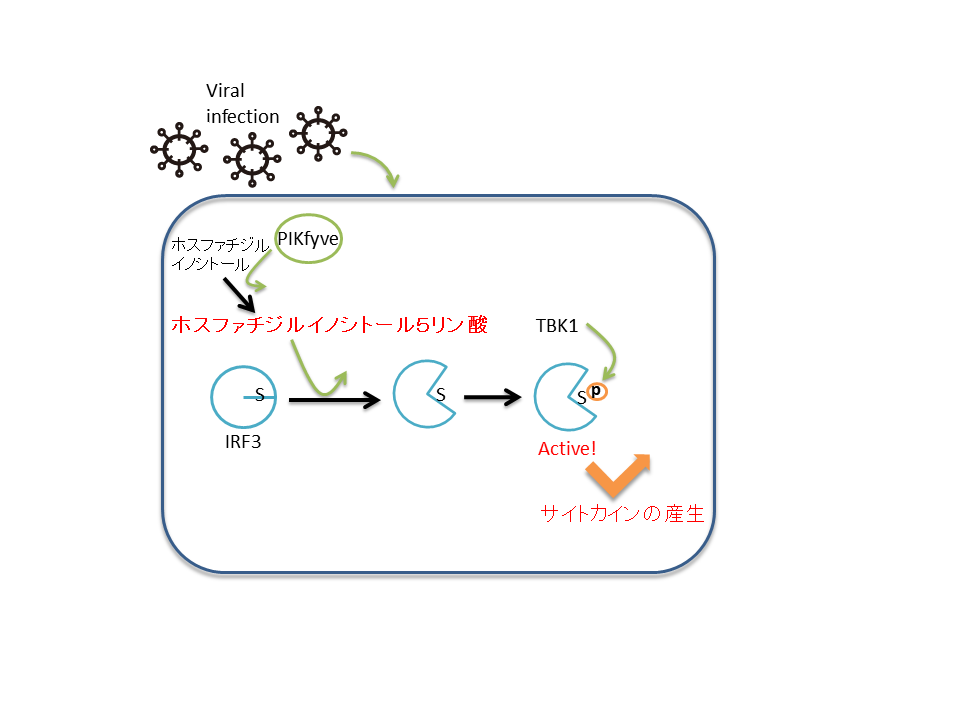
図3 ホスファチジルイノシトール5リン酸による自然免疫シグナルの制御メカニズム
ウイルスが感染するとホスファチジルイノシトールがPIKfyveによりリン酸化されホスファチジルイノシトール5リン酸が産生される。ホスファチジルイノシトール5リン酸はIRF3と結合することで、リン酸化サイトがTBK1により効率的にリン酸化されるようになる。
(2013年09月11日掲載)
この記事をシェア ![]()