Research 研究成果の紹介
植物の形態にひそむ周期性を変調させる仕組みを発見
魅力的な花き類の創出に期待
植物の形態にひそむ周期性を変調させる仕組みを発見
魅力的な花き類の創出に期待
【概要】
奈良先端科学技術大学院大学(学長:塩﨑一裕) 先端科学技術研究科 バイオサイエンス領域の池内桃子特任准教授と京都府立大学の爲重才覚講師(研究当時:名古屋大学トランスフォーマティブ生命分子研究所、 横浜市立大学、 奈良先端大を含む)、奈良先端大の土田岳志(研究当時:博士前期課程)らは、広島大学大学院統合生命科学研究科の藤本仰一教授、名古屋大学トランスフォーマティブ生命分子研究所/テキサス大学の鳥居啓子教授、名古屋大学遺伝子実験施設の打田直行教授、東京農工大学の笠原博幸教授、熊本大学の相田光宏教授らの研究グループと共同で、植物の形態形成の周期性を変調させる仕組みを発見しました。形態形成を司る植物ホルモンのオーキシン(注1)の空間的なパターンは、オーキシンの輸送によって創出されるという説が長年受け入れられてきましたが、パターンの周期性を決める仕組みはこれまで明らかになっていませんでした。今回の研究はEPFL2(EPIDERMAL PATTERNING FACTOR-LIKE 2)(注2)というペプチドホルモンとの相互抑制的な関係性が、オーキシンの周期的な空間パターンを変調させることを新たに見出しました。これは、従来モデルを20 年ぶりに書き換える重大な発見であるといえます。本研究の成果は、花卉(かき)類や園芸植物の形を操作する技術の創出につながることが期待できます。
この研究成果は、イギリスの学術誌「Nature Communications」オンライン版に2025 年11 月13 日(木)午後7 時(日本時間)に掲載されました(DOI:10.1038/s41467-025-65792-y)。
 【背景と目的】
【背景と目的】
ヒマワリやマツボックリなどのフィボナッチらせん(注3)に代表される植物の周期的な構造は、葉や花などの器官を一定間隔で繰り返し生み出すことで形作られています。また、ひとつひとつの器官の形状に着目すると、葉の縁には鋸歯(きょし)と呼ばれるギザギザ構造が一定の間隔で形成されていることがわかります(図1)。こうした器官や鋸歯等の突起構造は、植物ホルモンであるオーキシンが局所的に蓄積することによって形成されます。オーキシンの輸送体がオーキシン濃度の高い細胞の方へ流す働きがあることで自発的に周期的なパターンが生み出されることが、実験およびコンピュータシミュレーションの結果に基づいて20 年ほど前から提唱されていました1,2,3,4。この説によると、オーキシンの輸送と拡散の関係によって周期性が決まると想定されます。しかし、実際の植物において周期性が変わる観察例はなく、周期性が決まる仕組みは未解明でした。
 【研究手法と成果】
【研究手法と成果】
周期性を制御する仕組みを理解する手がかりは、EPFL2 と呼ばれるペプチドホルモンを作れない変異体の解析から得られました。鋸歯形成に先立って葉の縁に形成されるオーキシン蓄積部位を観察すると、EPFL2 の機能が損なわれたepfl2変異体ではその形成間隔が短くなっていることを発見しました(図2)。以前の研究によって、EPFL2 とオーキシンの間に相互に抑制し合う制御関係があることがわかっていました5。
そこで研究グループは、従来モデルのオーキシンと輸送体の制御関係にオーキシンとEPFL2 の相互抑制関係を組み込んだ新しいモデルを構築しました。葉を模した多細胞列をコンピュータ上で再現しEPFL2 がオーキシン蓄積部位の周期性に与える影響を調べた結果、EPFL2 の量が少なければ形成間隔が短くなるという観察結果を再現しました。さらに、EPFL2 の量を通常条件よりも増やせば間隔がもっと長くなり、一方でEPFL2 の量を極端に減らすとひとつひとつの蓄積部位が明瞭に形成されなくなるというシミュレーションの結果が得られました。研究グループは実験的にEPFL2 の量を改変した植物体を作り出し、これらのシミュレーション結果を裏付けることに成功しました。
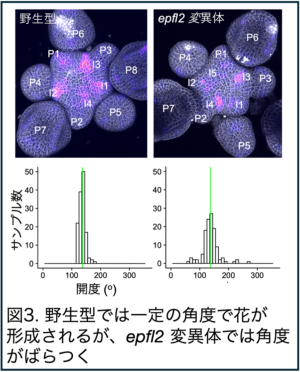
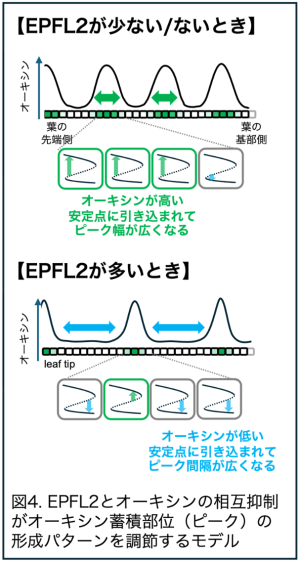
また、EPFL2 が規則的な器官配置パターンの形成に必要であることも見出し、EPFL2 は植物の形作りを一般的に制御する重要な因子であることを証明しました(図3)。こうして、実験とシミュレーションの両面のアプローチに基づき、EPFL2 とオーキシンの相互抑制的関係が繰り返し起こる形態形成イベントの周期性を変調させる仕組みを提案しました(図4)。
 【今後の展開】
【今後の展開】
輸送体によってオーキシン蓄積部位が周期的に生み出される仕組みは、チューリングパターン(注4)と呼ばれる一般的な規則正しいパターン形成と類似した仕組みと考えられています6。今回の研究は、相互抑制関係とチューリングパターンを組み合わせることで周期性を変調させるという、新しいパターン形成の仕組みを提案しています。植物の形態形成の制御に留まらず、自然界に見られる様々な周期的パターン形成においても同様の仕組みが働いている可能性が考えられます。また、サニーレタスのように複雑に入り組んだ葉の構造やカーネーションなど花弁の辺縁構造も鋸歯と共通した仕組みで作られるため、鑑賞用の花卉類や園芸植物の形質を改変して魅力的な形の植物を生み出す技術の開発に利用できる可能性があります。
【謝辞】
本研究は、科学技術振興機構(JST)創発的研究支援事業(JPMJFR214H)、日本学術振興会(JSPS)新学術領域研究「植物の周期と変調」(JP20H05431, JP22H04713)・新学術領域研究「植物多能性幹細胞」( JP17H06476 )・基盤研究B(JP26291057)・若手研究(JP20K15807)などの支援を受けて行われました。
【用語解説】
注1 オーキシン:植物の成長を調節する代表的な植物ホルモンの一つ。茎の伸長や根の形成、葉や花の配置など、体の各部の形づくり全般に深く関わっており、農業や研究目的で人為的に投与することも多い。
注2 EPFL2:EPIDERMAL PATTERNING FACTOR-LIKEs を含む一群のペプチド(EPF/EPFLs)のうちの一種。EPF/EPFLs は植物が体の各部の形成を調節するホルモン様のペプチド分子で、多くの植物が持っている。EPFL2 は特に葉の形態、胚珠(種子になる器官)の形成、分裂組織のサイズなどに関わることが知られている。
注3 フィボナッチらせん:らせんの列の数がフィボナッチ数列(前の2 つの数を足した数が次の数になるという規則性を持つ数列)に従う規則的な配置。
注4 チューリングパターン:物質の反応と拡散の仕組みによって自然に生じる模様、およびその理論。イギリスの数学者で暗号解読の功績やコンピュータ科学の創始者としても知られるアラン・チューリングが提唱した。動物の斑点や縞模様、植物の模様や形態などを説明する理論として知られる。
【引用文献】
1. Jönsson H, Heisler MG, Shapiro BE, Meyerowitz EM & Mjolsness E. An auxin-driven polarized transport model for phyllotaxis. Proc Natl Acad Sci U S A. 103, 1633-1638 (2006).
2. de Reuille, P. B. et al. Computer simulations reveal properties of the cell–cell signaling network at the shoot apex in Arabidopsis. Proc. Natl. Acad. Sci. USA. 103, 1627-1632. (2006).
3. Smith, R. S. et al. A plausible model of phyllotaxis. Proc. Natl. Acad. Sci. USA 103, 1301-1306. (2006).
4. Bilsborough GD et al. Model for the regulation of Arabidopsis thaliana leaf margin development. Proc Natl Acad Sci U S A. 108, 3424-9 (2011).
5. Tameshige T et al. A secreted peptide and its receptors shape the auxin response pattern and leaf margin morphogenesis. Curr Biol. 26, 2478-2485 (2016).
6. Sahlin P., Söderberg B. & Jönsson H. Regulated transport as a mechanism for pattern generation: capabilities for phyllotaxis and beyond. J. Theor Biol. 258, 60-70 (2009).
【掲載論文】
タイトル:Mutual inhibition between EPFL2 and auxin extends the intervals of periodic leaf morphogenesis(EPFL2 とオーキシンの相互抑制が葉の周期的な形態形成の間隔を伸ばしている)
著者:Toshiaki Tameshige#, Takeshi Tsuchida#, Yuuki Matsushita, Yuki Doll, Kaisei Maruyama, Takemoto Agui, Mitsuhiro Aida, Hiroyuki Kasahara, Keiko U Torii, Naoyuki Uchida, Koichi Fujimoto, Momoko Ikeuchi*
# 共筆頭著者
* 責任著者
掲載誌:Nature Communications
DOI:10.1038/s41467-025-65792-y
池内桃子特任准教授のコメント
今回の研究は、研究員の爲重さん(画像左)と修士課程に在籍していた土田さん(画像中央)が中心的な役割を果たし、数理生物学やホルモン分析など様々な専門家と共同研究を展開することで大きな成果につながりました。筆頭著者の二人が栄転と卒業でラボを離れた後にも、ラボメンバーが実験や解析を引き継いで研究を完成させられて良かったです。本学のLiSCO が誇る顕微鏡のおかげで、優れたイメージング解析が実現しました。また、オープンアクセス雑誌への掲載費用をJ PEAKS 事業からご支援いただきまして誠にありがとうございました。

【植物再生学研究室】
研究室紹介ページ:https://bsw3.naist.jp/courses/courses117.html
研究室ホームページ:https://bsw3.naist.jp/ikeuchi/
(2025年11月14日掲載)
この記事をシェア ![]()