Research 研究成果の紹介
世界初!植物の幹や根が太る側方成長を制御する
巧妙な仕組みを解明
~作物や樹木の成長強化、収量の向上に期待~
世界初!植物の幹や根が太る側方成長を制御する
巧妙な仕組みを解明
~作物や樹木の成長強化、収量の向上に期待~
【概要】
奈良先端科学技術大学院大学(学長:横矢直和、奈良県生駒市)先端科学技術研究科 植物発生シグナル研究室の宮島 俊介(みやしま しゅんすけ)助教、中島 敬二(なかじま けいじ)教授、フィンランド Helsinki大学、英 セインズベリー研究所らの国際共同研究グループは、樹木の幹が太くなるなど植物の肥厚(側方成長)の仕組みの中で、肥厚の出発点である前形成層での細胞分裂を活性化するPEAR遺伝子群を世界で初めて発見しました。また、植物はPEAR遺伝子の機能によって、様々な細胞同士の情報のやり取りを統合し、側方成長を駆動する細胞分裂活性を空間的に調節していることも世界で初めて明らかにしました。
植物の成長は、地上および地下方向へ伸びてゆく「先端成長」と、茎や根などを放射方向(横方向)に肥厚させる「側方成長」に分けられます。この側方成長は、植物の茎や根の維管束組織の中の「前形成層・形成層」と呼ばれる細胞集団が活発に細胞分裂することで木部や師部といった維管束組織を肥大化させ、結果的に茎や根を太らせるプロセスであることが古くから知られていました。しかしながら、この前形成層・形成層の「どこで」、「どのように」細胞分裂が起きるのか、またそれら細胞分裂を制御する仕組みが何であるかは、全く未解明でした。
宮島助教らの国際共同研究グループは、維管束植物のモデルであるシロイヌナズナ (Arabidopsis thaliana)の若い根の前形成層(根の先端にある形成層)を側方成長のモデル系に用い、まず、側方成長における細胞分裂の空間的な分布を調べました。その結果、前形成層での細胞分裂が、師部細胞とその周囲の細胞群に集中していることを見出しました。
研究グループは、この側方成長を制御する分子機構を解明するために、師部細胞で特異的に発現するPEAR遺伝子群に着目しました。PEAR遺伝子群は、他の遺伝子群の働きを制御する植物に特有のDOF型転写因子タンパク質をコードしています。実験の結果、PEARタンパク質は、師部細胞で作られたあと、原形質連絡という細胞間を繋ぐトンネルを通って細胞間を移動し、活発に分裂する細胞群に蓄積することを見出しました。次いで、PEAR遺伝子群を多重破壊した変異体(pear変異体)では、前形成層の細胞分裂が起こらず、逆にPEARタンパク質の量を過剰にすると、前形成層に過剰な細胞分裂を引き起こすことを発見しました。さらに、実験を重ねたところ、PEARタンパク質が、植物ホルモンや低分子量RNAを介した細胞間の情報のやり取りを制御し、根の前形成層の細胞分裂を空間的に統御する鍵因子であることを突き止めました。
本研究は、維管束植物を太らせる側方成長のしくみを分子レベルで解明した世界初の成果です。植物の側方成長を自在に操作する技術の確立を通じ、根菜類をはじめとした農作物の収量増加やバイオマス生産効率の向上など、食糧やエネルギーの安定供給へ貢献も期待されます。
この研究成果は、英国時間の2019年1月9日 (水) 午後6時 【プレス解禁日時:日本時間平成31年1月10日(木) 午前3時】付で、Natureのオンライン版に発表されました。
【論文情報】
タイトル:
Mobile PEAR transcription factors integrate positional cues to prime cambial growth
(細胞間を移行するPEAR転写因子は細胞の位置情報を統合することで側方成長を開始させる。)
著者:
Shunsuke Miyashima1,2,15, Pawel Roszak1,3,15, Iris Sevilem1,15, Koichi Toyokura3,4, Bernhard Blob3, Jung-ok Heo1,3, Nathan Mellor5, Hanna Help-Rinta-Rahko1 , Sofia Otero3, Wouter Smet6,7,8, Mark Boekschoten9, Guido Hooiveld9, Kayo Hashimoto2,10, Ondřej Smetana1 , Riccardo Siligato1 , Eva-Sophie Wallner11, Ari Pekka Mähönen1 , Yuki Kondo12, Charles W. Melnyk3,13, Thomas Greb11, Keiji Nakajima2 , Rosangela Sozzani14, Anthony Bishopp5, Bert De Rybel6,7,8* & Ykä Helariutta1,3*
1Institute of Biotechnology, HiLIFE/Organismal and Evolutionary Biology Research Programme, Faculty of Biological and Environmental Sciences, Viikki Plant Science Centre, University of Helsinki, Helsinki, Finland. (フィンランド ヘルシンキ大学)
2Graduate School of Science and Technology, Nara Institute of Science and Technology, Nara, Japan.(奈良先端科学技術大学院大学)
3The Sainsbury Laboratory, University of Cambridge, Cambridge, UK. (イギリス、セインズベリー研究所 ケンブリッジ大学)
4Department of Biological Sciences, Graduate School of Science, Osaka University, Toyonaka, Japan.(大阪大学)
5Centre for Plant Integrative Biology (CPIB) and School of Biosciences, University of Nottingham, Nottingham, UK. (イギリス、ノッティンガム大学)
6Department of Plant Biotechnology and Bioinformatics, Ghent University, Ghent, Belgium. (ベルギー ゲント大学)
7VIB Center for Plant Systems Biology, Ghent, Belgium. (ベルギー VIB)
8Laboratory of Biochemistry, Wageningen University, Wageningen, The Netherlands.
9Nutrition, Metabolism and Genomics Group, Division of Human Nutrition, Wageningen University, Wageningen, The Netherlands. (オランダ ヴァーハニンゲン大学)
10Graduate School of Humanities and Sciences, Nara Women's University, Nara, Japan. (奈良女子大学)
11Centre for Organismal Studies (COS), Heidelberg University, Heidelberg, Germany. (ドイツ ハイデルブルグ大学)
12Department of Biological Sciences, Graduate School of Science, The University of Tokyo, Tokyo, Japan. (東京大学)
13Department of Plant Biology, Swedish University of Agricultural Sciences, Uppsala, Sweden.(スウェーデン スウェーデン農業大学)
14Department of Plant and Microbial Biology, North Carolina State University, Raleigh, NC, USA. (アメリカ ノースカロライナ大学)
15共同第1著者 *共同責任著者
掲載誌:
Nature
DOI:
10.1038/s41586-018-0839-y
掲載日時:
2019年1月9日 オンライン版
【背景】
植物の成長は、地上や地下方向など縦に伸びる「先端成長」と、茎や根などを放射方向(横方向)に肥厚させる「側方成長」に分けられます。側方成長は、植物の茎や根の維管束組織にある「形成層*1」や若い根や茎の先端にある「前形成層*1」と呼ばれる細胞群が活発に分裂し、新たに作られた細胞が木部や師部といった組織に分化して維管束組織を肥大化させることで、茎や根などを太らせるプロセスです。先端成長が発芽直後から継続するのに対し、側方成長は、若い組織に存在する前形成層が一過的に成長した後一旦成長を停止し、器官発生の後になって再び活性化するという段階的なパターンを示します。
側方成長では、植物ホルモンの一種であるサイトカイニン*2が、前形成層および形成層での細胞分裂を活性化する事が知られていました。しかしながら、サイトカイニンがどのような遺伝子を活性化して細胞分裂を駆動するかは全く分かっていませんでした。また、前形成層や形成層を構成する様々な細胞のうち、どの細胞がどのように分裂して側方成長を生み出しているのかも、ほとんど知られていませんでした。
宮島助教らの国際研究グループは、側方生長の仕組みを明らかにするために、モデル被子植物のシロイヌナズナ(Arabidopsis thaliana) *3の若い根の先端部に存在する前形成層に着目しました。その結果、根の前形成層の側方成長では、おもに将来師部に分化する師部細胞とそれらに隣接する細胞群が分裂すること、また、サイトカイニンの作用を通じて師部細胞で作られるPEARタンパク質が、師部細胞からその周辺の細胞に移動することで、前形成層での細胞分裂を促進していることを明らかにしました。さらに、PEARタンパク質は、前形成層で働く様々な細胞間情報伝達物質の働きを統合し、「分裂する細胞」と「分裂しない細胞」を正確に決定していることを発見しました。維管束植物の成長制御については、茎や根の先端を伸長させる頂端部分裂組織の活性化について多くの研究がなされていましたが、植物を太らせバイオマスを増加させる側方成長についての理解は遅れていました。本研究は、維管束植物の側方成長の根幹をなす原理を分子レベルで解明した世界初の成果となります。
【実験の結果】
シロイヌナズナの根の前形成層の側方成長では、その先端部にある始原細胞と呼ばれる数個の細胞群が細胞分裂を繰り返し、これによって生じた新たな細胞群が、師部や木部といった組織を構築することが知られていました(図1)。研究グループは、まずこれらの細胞分裂が、「どこでどのように起こっているのか」を明らかにするために、前形成層での細胞分裂を詳細に観察しました。その結果、将来師部に分化する細胞(師部細胞)とその側方に隣接する細胞が、集中的に細胞分裂を繰り返していることを明らかにしました。同時に、ほとんど細胞分裂をしない細胞群の存在も見出しました。これらの結果から、前形成層での細胞分裂活性は細胞の「位置」に依存する、という法則を見出しました(図1)。
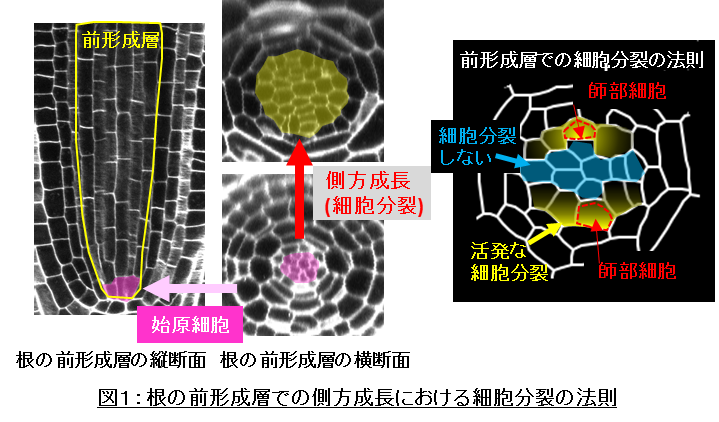
師部細胞とその側方の細胞で集中的な細胞分裂が観察されたことから、研究グループは、これらの細胞群に、「細胞分裂を活性化する分子」が存在するのではないかと予想しました。シロイヌナズナをはじめとした植物においては、遺伝子の発現に関わる転写因子*4と呼ばれるタンパク質が、原形質連絡*5とよばれる細胞と細胞を繋ぐ通路を介して、細胞間を移動することで「位置に応じた細胞の機能」を制御することが広く知られています。そこで、師部細胞で特異的に発現する6つのDOF型転写因子*6に着目し、これらをPHLOEM EARLY DOF (PEAR)転写因子群と名付けて機能解析を進めました。解析の結果、PEARタンパク質は、師部細胞で作られたのち、原形質連絡を通じて師部細胞の周辺細胞に移動することを見出しました(図2)。また、6つのPEAR遺伝子すべてが機能しない植物体(pear変異体)では、前形成層の細胞分裂が起こらず(図3)、逆に、PEAR遺伝子の発現量を過剰にした植物体(PEAR過剰発現体)では、前形成層の細胞分裂が過剰に起こることを見出しました(図3)。さらに、PEAR遺伝子は、側方成長の細胞分裂を促すことが知られていたサイトカイニンの作用を通じて活性化されることも見出しました。これらの結果から、サイトカイニンの機能によって作られるPEARタンパク質が、原形質連絡を介して細胞をまたいで分配されることで、師部細胞付近で集中的な細胞分裂が起こることが明らかになりました(図3)。

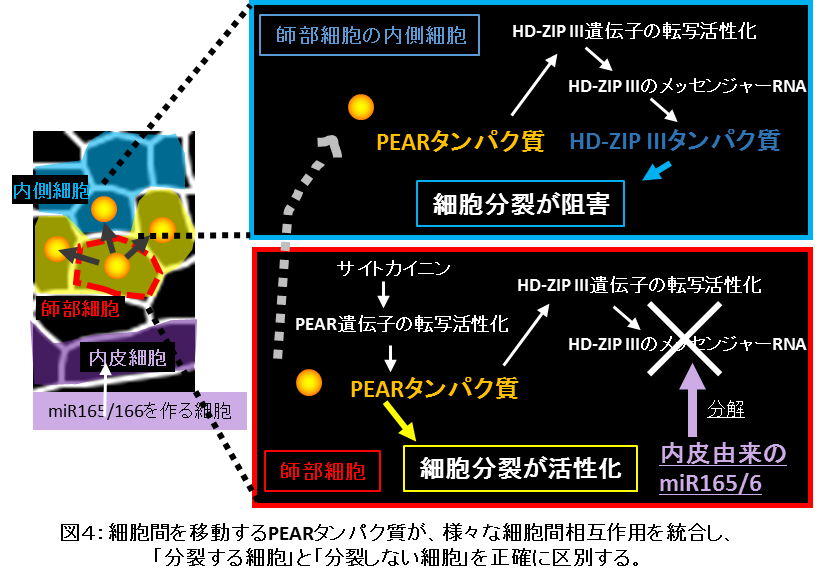
一方で、師部細胞に根の内側(中央側)で接する細胞(内側細胞)は、PEARタンパク質が存在するにも関わらず分裂していません(図4)。このことから、内側細胞にはPEARタンパク質の機能を阻害する仕組みがあると考えました。そこで、研究グループでは、PEARタンパク質とは異なる転写因子であるホメオドメイン型転写因子*7というタイプのクラスIII HD-ZIP (HD-ZIP III)群に着目しました。同研究グループは、HD-ZIP III転写因子群は、前形成層での細胞分裂を抑制することを以前に報告していました。PEARタンパク質とHD-ZIP IIIタンパク質との関係性を解析した結果、内側細胞に移動したPEARタンパク質は、HD-ZIP III遺伝子の転写を活性化し、HD-ZIP IIIタンパク質の量を増大させることで、細胞分裂を阻害することを発見しました(図4)。師部の側方にある細胞においてはHD-ZIP IIIによる分裂阻害がおこりませんが、これは前形成層の外側にある内皮細胞で作られたマイクロRNA*8(miR165/166)が、HD-ZIP III遺伝子の発現を阻害しているためであることも見出しました。このような複雑かつ巧妙な分子機構を通じ、PEARタンパクが前形成層の中で「分裂する細胞」と「分裂しない細胞」を正確に指定していることを明らかにしました(図4)。
【今後の発展】
本研究により、維管束植物の側方成長において細胞分裂を空間的に統御する分子ネットワークが発見されました。本研究でその機能が発見されたPEARタンパク質群は、この分子ネットワークで中心的な機能を果たしています。植物はPEARタンパク質の機能を通じて様々な細胞間情報伝達を統合し、前形成層のなかで「分裂する細胞」と「分裂しない細胞」を正確に区別化していることが明らかになってきました。本研究成果は、植物の成長を遺伝子・分子レベルで解明する学術研究の展開において基盤となる作動原理を提示したものです。
樹木の幹やサツマイモなどの根菜作物の肥大化は、側方成長の著しい進行により達成されています。本研究で明らかになった遺伝子群や情報伝達物質は、維管束植物に共通して存在することが知られていることから、本成果を基盤とすることで、樹木や農作物などの有用植物の肥大化を自由自在にコントロールする技術の開発といった応用研究が展開することが期待されます。
【用語説明】
1.前形成層、形成層
維管束植物の維管束組織に存在する細胞群。細胞分裂をすることで、あらたな細胞を生みだす。生じた細胞は、水を通す木部(導管)や光合成産物である糖などを輸送する師部(師管)に分化する。
2.サイトカイニン
古典的に知られた植物ホルモンの一種である低分子有機化合物。一般的に、植物の細胞分裂を促進し、カルスからの葉や茎などの地上部組織の誘導効果を持つ。
3.シロイヌナズナ
初めて全ゲノム配列が解読された維管束植物で、アブラナ科の1年草。遺伝学的解析や組織形成のイメージングに適したモデル植物として、世界的に広く利用されている。
4.転写因子
ゲノムDNAの調節領域に結合し、mRNAへの転写を調節するタンパク質。転写を活性化する因子と抑制する因子がある。シロイヌナズナのゲノムには約2000個の転写因子がコードされている。
5.原形質連絡(げんけいしつれんらく、Plasmodesmata (プラズモデスマータ))
植物細胞において、隣接する細胞の細胞壁を貫通して作られるトンネル状の通路。隣接した細胞間で細胞質を直接連結するため、細胞膜に遮られることなく分子をやりとりすることに用いられる。原形質連絡を通じて、ショ糖のような栄養源といった低分子物質のみならず、タンパク質や低分子RNAなどの制御因子が選択的・非選択的に移動する。全ての陸上植物に存在する。動物細胞にはギャップ結合という進化的な由来が全く異なる相同な細胞構造が存在する。
6.DOF型転写因子
植物特有の転写因子群であり、シロイヌナズナのゲノムには36種類コードされている。
7.ホメオドメイン型転写因子
転写因子の一種で、動物と植物に共通して存在する。動植物を通じて、様々なホメオドメイン型転写因子が、個体の形作りに機能していることが知られている。
8.マイクロRNA
21-24塩基からなる一本鎖RNA。通常の遺伝子のメッセンジャーRNA(mRNA)と同様に、ゲノムDNAから転写されたRNAから作られるが、通常のmRNAと異なりタンパク質をコードせず、21-24塩基の短い機能性RNAに切り詰められたのちに、相同な配列を持つ他の遺伝子のmRNAの分解や翻訳阻害を通じて、それらの機能を阻害する。
[植物発生シグナル研究室]
研究室紹介ホームページ:http://bsw3.naist.jp/courses/courses110.html
研究室ホームページ:http://bsw3.naist.jp/nakajima/
(2019年02月05日掲載)
この記事をシェア ![]()