Research 研究成果の紹介
植物で生殖細胞をつくり出す主役の遺伝子を発見 ~生殖機構の解明や、繁殖・育種技術への応用に期待~
バイオサイエンス研究科、植物発生シグナル研究室の中島敬二教授らの研究グループは、京都大学、大阪市立大学、広島大学、神戸大学、近畿大学との共同研究により、植物において生殖細胞の形成や多能性の発現を制御する遺伝子を発見しました。植物の生殖細胞形成を中心的に制御する遺伝子の発見は初めてのことです。今後はこの遺伝子を手掛かりに、植物が生殖細胞を作るメカニズムや、生殖細胞が分化多能性を獲得する仕組みを明らかにできると期待されます。
この研究成果をまとめた論文は、平成28年6月24日(金)にCurrent Biologyのオンライン版に掲載され、朝日新聞に記事として掲載されました。
プレスリリース詳細 ( 大学HP http://www.naist.jp/ 内コンテンツ )
中島敬二教授による研究成果の解説
 有性生殖は、両親の遺伝情報を混ぜ合わせて子孫の形質に多様性をもたらす プロセスで、生物進化の原動力です。有性生殖には、卵と精子という多能性と受精能を併せ持った特殊な細胞を作る必要があります。動物の生殖細胞系列は初期胚の段階で既に取り分けられていますが、植物の生殖細胞は個体発生のずっと後に花という生殖器官の中に作られます。このため植物の生殖細胞形成では体細胞を初期化(リプログラミング)し、さらに受精に適した形態へ変化させる必要があります。植物の生殖細胞形成において、このような複雑な過程が制御される仕組みは未解明です。
有性生殖は、両親の遺伝情報を混ぜ合わせて子孫の形質に多様性をもたらす プロセスで、生物進化の原動力です。有性生殖には、卵と精子という多能性と受精能を併せ持った特殊な細胞を作る必要があります。動物の生殖細胞系列は初期胚の段階で既に取り分けられていますが、植物の生殖細胞は個体発生のずっと後に花という生殖器官の中に作られます。このため植物の生殖細胞形成では体細胞を初期化(リプログラミング)し、さらに受精に適した形態へ変化させる必要があります。植物の生殖細胞形成において、このような複雑な過程が制御される仕組みは未解明です。
今から5年ほど前に、私たちの研究グループは、シロイヌナズナのRKD4遺伝子が細胞初期化能をもつことを明らかにしました。RKD4を発芽後の幼植物で人工的に過剰発現させると、葉や根から初期化された細胞の塊が作られ、そこから大量の胚が作られます。RKD4は、本来は初期胚で特異的に発現しており、ノックアウトすると胚発生が正常に進まなくなります。
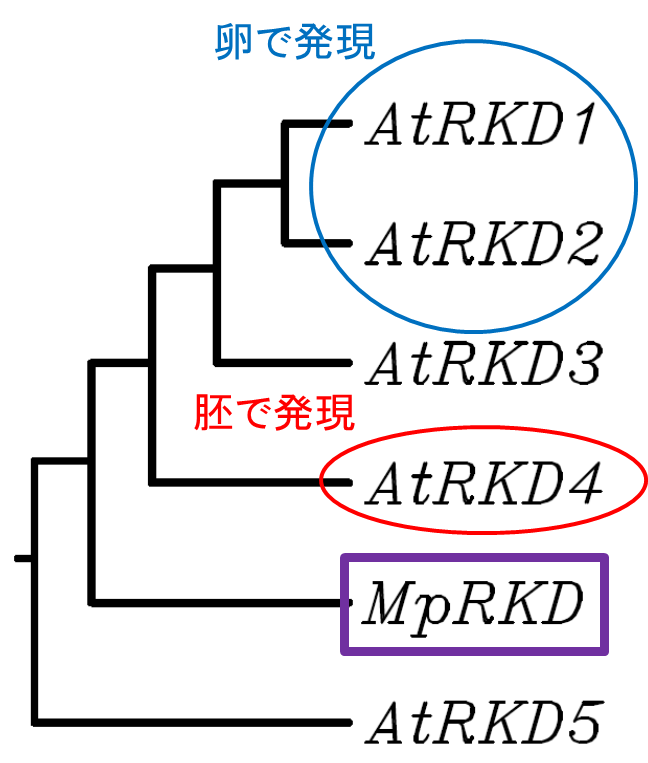
RKD4は胚発生の制御因子でしたが、細胞の初期化は胚発生に先立つ卵や花粉の形成時に起こっています。シロイヌナズナのゲノムにはRKD1-RKD5の合計5つのRKD遺伝子があり、これらのうちの少なくともRKD1とRKD2はRKD4と同様に初期化能をもち、卵細胞で強く発現していることが分かっていました。つまり、RKDはもともと生殖細胞の形成因子であり、RKD4だけが進化の過程で胚発生を制御するようになったのではないかと考えました。
しかし、シロイヌナズナなどの種子植物でこの仮説を証明するのは容易ではありません。その理由は、種子植物にはRKD遺伝子が複数あり重複した機能を持っている可能性が高いこと、半数体である生殖細胞では必須遺伝子を壊すことができないこと、さらに種子植物の生殖細胞がつぼみの奥深くで短時間のうちに作られるため、その形成過程を観察するのが非常に難しいためです。植物の発生原理の概要は過去20年の間にほぼ解明されましたが、生殖細胞の形成機構についてはほとんど未解明のままです。その理由は、広く用いられているモデル植物が生殖細胞の研究には不向きなためです。
そこで私たちは陸上植物の祖先であるゼニゴケに注目しました。ゼニゴケは形質転換系と遺伝子を自在に破壊する技術が確立されており、新しいモデル植物として急速に普及しています。さらに全ゲノムがほぼ解読され、非常に単純なゲノム構成をもつこと、つまり機能重複した遺伝子をほとんど持たないことが分かっています。調べてみると、ゼニゴケのゲノムにはRKD遺伝子が1つしかありませんでした。そこでこのRKD遺伝子を、ゼニゴケの学名(Marchantia polymorpha)の頭文字をとってMpRKDと名付けました(図1)。

ゼニゴケは雌雄異株の半数体植物で、メスの造卵器に卵細胞が、オスの造精器に精子が作られます(図2)。まずMpRKDがどこで発現しているのか調べたところ、メスでは成熟過程の卵細胞、オスでは精子のもとになる精原細胞で発現していました。一方で受精後の接合子では発現していませんでした。このことからMpRKDはシロイヌナズナのRKD1やRKD2と同様に、受精前に働いており、受精後に働くRKD4とは異なっていることが分かりました。
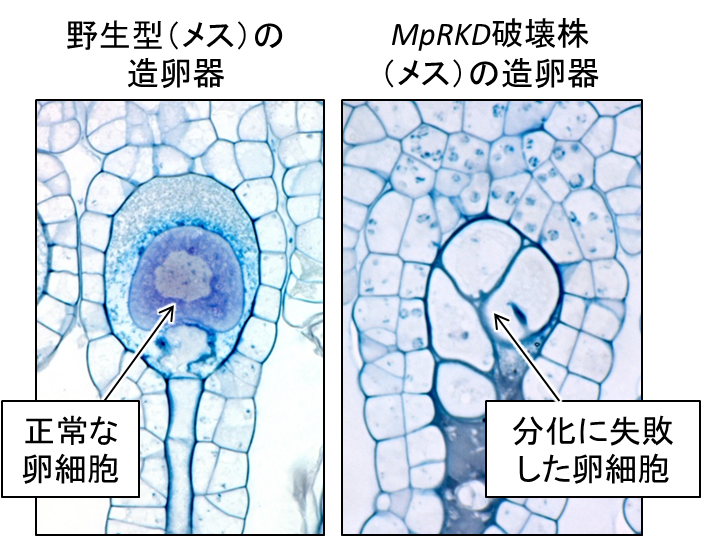
次にMpRKDの破壊株を作成しました。ゼニゴケの破壊株は、生殖細胞を経ずに作ることができます。メスの破壊株では、卵細胞の前駆細胞までは問題なく作られるものの、その後の卵に特有な分化がおこらず、異常な細胞分裂と液胞化を経て崩壊していました(図3)。また破壊株の卵は精子を誘引できませんでした。
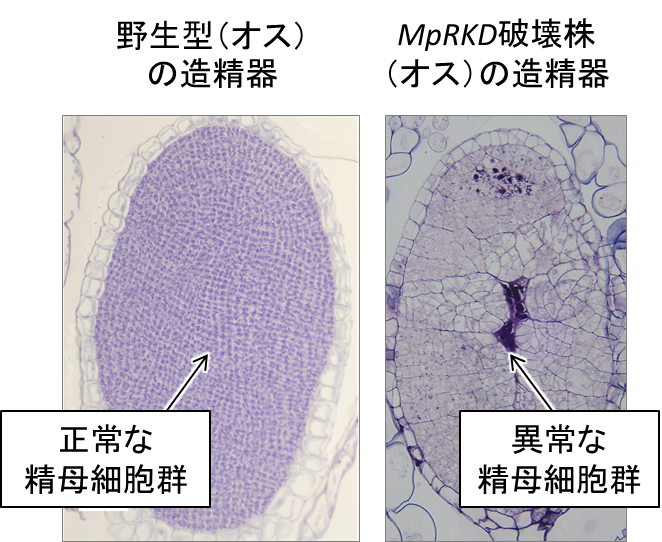
またオスの破壊株では、造精器までは正常に作られるものの、その内部にある精原細胞が同調的に分裂せず、正常な精子形成が起こりませんでした(図4)。一方で生殖細胞以外には顕著な異常がありませんでした。
以上の結果から、RKD機能の進化的な起源は、生殖細胞の分化や初期化の制御であると結論づけました。種子植物に至るまでの進化の過程で遺伝子の数が増え、シロイヌナズナではそのうちの1つであるRKD4が、受精後の胚発生を制御するようになったと考えられます。これを証明するには、進化系統樹でコケと種子植物の間に位置するシダなどを用いて解析することが必要です。
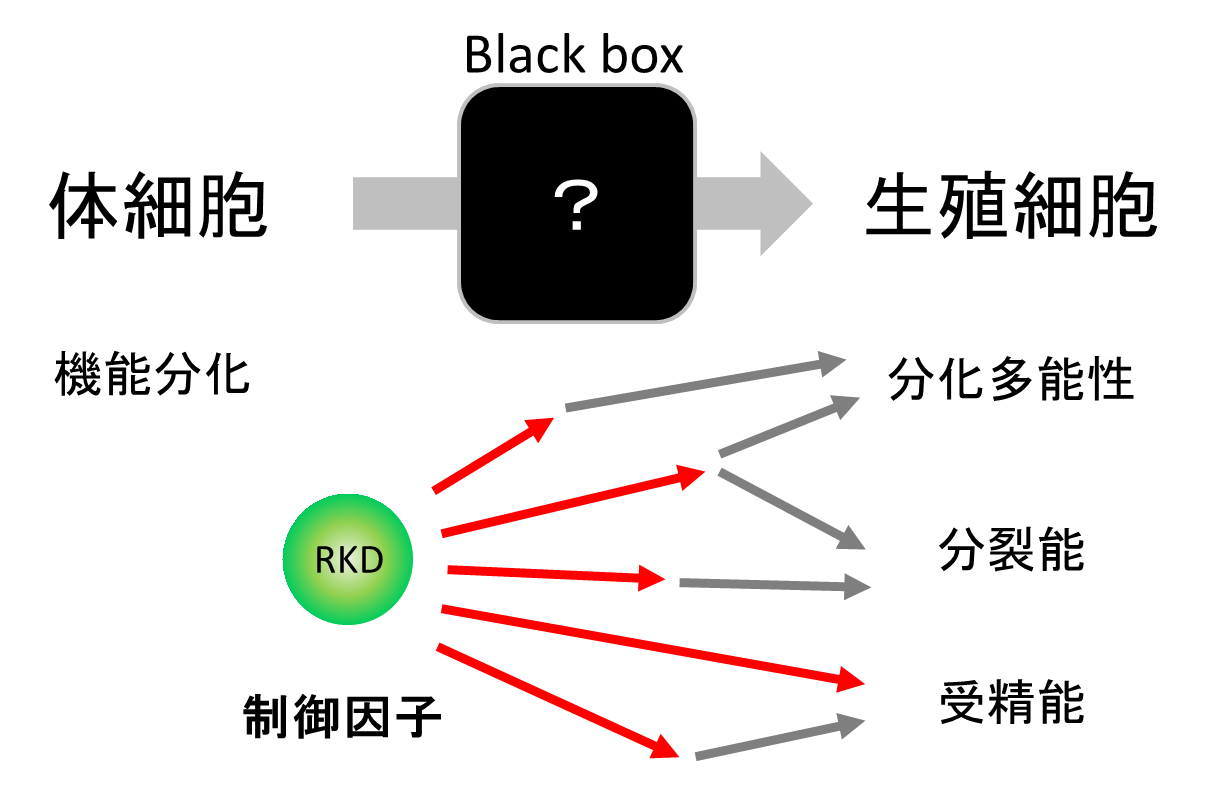
RKDが生殖細胞形成の鍵を握る制御因子であることが明らかになり、今後はRKDを起点に生殖細胞の分化機構を明らかにできると期待されます(図5)。またRKDを用いて試験管内で生殖細胞を誘導できれば、作物や花卉の育種が加速するかもしれません。この研究成果は、植物における生殖細胞の制御因子を明らかにしたのみならず、ゼニゴケという新しいモデル植物をうまく使うことで、既存のモデル植物の困難を克服したことも特筆すべき点です。
この研究は、私の研究室に所属していた厚井聡博士(現、大阪市立大学講師)が中心になって進めたもので、研究室の他の研究員や大学院生、テクニシャンらも参画しています。またゼニゴケの遺伝学や顕微鏡観察については、京都大学の河内孝之教授をはじめ、広島大、神戸大、近畿大のエキスパートの先生方と共同でデータを取得しました。共同研究者の皆様には、この場を借りてお礼を申し上げます。またこの研究は、平成23年度 JSTさきがけ採択課題「植物生産能の高度利用に向けた「植物iPS遺伝子」の応用展開」、科学研究費補助金、新学術領域研究「植物発生ロジック」および基盤研究(B)、倉田奨励金の支援を受けて行われました。
【論文情報】
タイトル:
An evolutionarily conserved plant RKD factor controls germ cell differentiation
著者:
Satoshi Koi, Tetsuya Hisanaga, Katsutoshi Sato, Masaki Shimamura, Katsuyuki T. Yamato, Kimitsune Ishizaki, Takayuki Kohchi and Keiji Nakajima
Current Biology 26, July 11, 2016
DOI: http://dx.doi.org/10.1016/j.cub.2016.05.013
【植物発生シグナル】
研究室紹介ページ:http://bsw3.naist.jp/courses/courses110.html
研究室ホームページ:http://bsw3.naist.jp/nakajima/
(2016年06月27日掲載)
この記事をシェア ![]()