Research 研究成果の紹介
植物の自殖抑制(多様性保持)に関わる新たな因子
種内の遺伝的多様性を保持することは、生物が変動する環境を生き抜く上で極めて重要であるが、高等植物では自家不和合性と呼ばれる機構がその役割の一端を担っている。これは、植物が自己由来の花粉と非自己由来の花粉を識別し、自己花粉の発芽・伸長を選択的に阻害することで、遺伝的多様性を失わせる原因となる「自殖(自家受精)」を防ぐ仕組みである。我々はアブラナ科植物を材料に研究を進め、雌ずい先端の乳頭細胞に存在する受容体型キナーゼSRK(S-locus protein kinase)が、花粉表層の自己マーカータンパク質SP11(S-locus protein 11)を受容することによって、自己の花粉を識別していることを明らかにしてきた。また、自家和合性の変異株の解析から、乳頭細胞内に存在する細胞質キナーゼMLPK(M-locus protein kinase)が自家不和合反応に関与することを明らかにしてきた。キナーゼとしての機能を欠損した変異型MLPKを持つ植物体は自己の花粉を拒絶できないことから、MLPKは自家不和合性の情報伝達系において重要な役割を果たしていると考えられるが、この分子の具体的な機能については明らかとされていなかった。
今回我々は、MLPK遺伝子からN末端の異なる2種類の蛋白質(MLPKf1、MLPKf2)が作られること、いずれも乳頭細胞内に存在するが、各々単独で自家不和合性情報を伝達する能力を持つことを明らかにした。また、MLPKf1とMLPKf2は、それぞれ異なる機構で細胞膜に結合していること、この細胞膜局在性が自家不和合性の情報伝達機能に必須であることを明らかにした。さらに、BiFC法(Bimolecular Fluorescent Complementation)による相互作用解析により、MLPKとSRKが細胞膜上で直接相互作用していることを明らかにし、MLPKがSRKと共に自己マーカータンパク質SP11に対する受容体複合体を形成しているという新たな分子モデル(図)を提唱した。
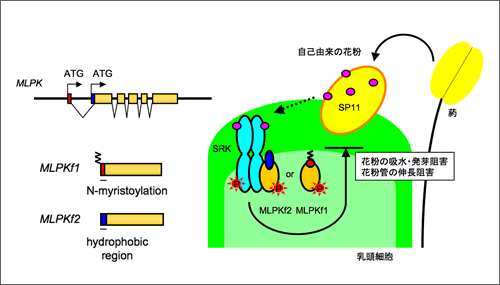
アブラナ科植物が自己花粉を認識する仕組み
MLPKはN末端領域の異なる2つの翻訳産物(MLPKf1、MLPKf2)を産生する。MLPKf1はN-ミリストイル化を介して、MLPKf2はN末端疎水性領域を介してとそれぞれ異なる機構で乳頭細胞の細胞膜に結合しており、SRKと受容体複合体を形成している。このSRK-MLPK受容体複合体が自己花粉由来のSP11による不和合シグナルを下流に伝達することで、花粉の吸水・発芽阻害、花粉管の伸長阻害といった自家不和合反応が引き起こされる。
掲載論文
Kakita, M., Murase, K., Iwano, M., Matsumoto, T., Watanabe, M., Shiba, H., Isogai, A., and Takayama, S. (2007) Two distinct forms of M-locus protein kinase localize to the plasma membrane and interact directly with S-locus receptor kinase to transduce self-incompatibility signaling in Brassica rapa. Plant Cell in press (Epub 2007 Dec 7; 10.1105/tpc.106.049999)
(2008年01月10日掲載)
この記事をシェア ![]()