

NEWS & TOPICS
- 2026.2A paper of Asako Katsuma (Dr. student) about L1-CAM-mediated glioblastoma invasion was published in Cancer Science.
- 2025.8A paper describing the mechanism of rapid leukocyte migration and gllioblastoma invation was published in Advanced Science. (EurekAlert!)(MedicalExpress)
- 2024.12A paper of Assistant Prof. Takunori Minegishi (about mechanical regulation of neuronal migration) was published in EMBO Journal.
- 2024.9Prof. Inagaki gave an invited talk at 25th Meeting of the International Society for Developmental Neuroscience.
- 2024.5Prof. Inagaki gave an invited talk at 6th Tri-lateral NAIST-TLL-CU Joint Symposium.
- 2024.2A preprint of D3 student Kio Yagami (about actin waves) was published in bioRxiv.
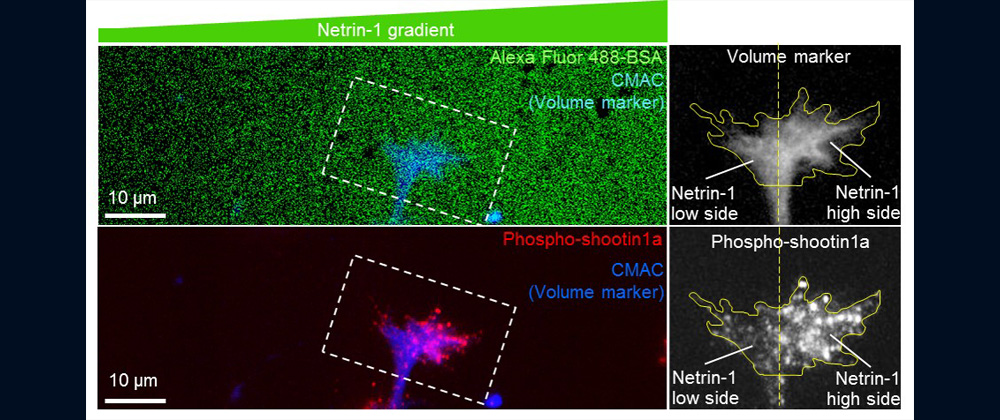
- 2024.2A paper of D3 student Qiu Zhen (about mechanical regulation of netrin-1-mediated axon guidance) was published on line in Frontiers in Molecular Neuroscience.
- 2023.7Prof. Inagaki and Assistant Prof. Minegishi gave a JoVE Webinar "Imaging techniques to analyze molecular mechanics for cell migration and axon guidance".(See here)
- 2023.3A paper of Ria Kastian (about axon guidance mediated by shootin1 dephosphorylation ) was published on line in Journal of Biological Chemistry.
- 2023.3Prof. Inagaki gave an invited talk at International Symposium on Mechanobiology for Human Health
- 2022.6A review of Takunori Minegishi (concerning mechanical regulation of synapse formation and plasticity) was published on line in Sem. Cell Dev. Biol.
- 2021.10Prof. Inagaki gave an invited talk at OIST Axonal Degeneration and Regeneration Workshop
- 2021.10A paper of Takunori Minegishi (concerning mechanobiological analyses of growth cone migration) was published in J. Vis. Exp.
- 2021.10A paper of Ria Kastian (concerning mechanobiological analyses of synapse formation) was published in Star Protoc.
- 2021.8A study of Kouki Abe (about the mechanism of mechanosensitive axon outgrowth) was published in Biophysical Journal.
- 2021.5A study of Ria Kastian (about shootin1-mediated mechanism for synaptic plasticity) was published in Cell Reports.
- 2020.11Ria Kastian gave an invited talk in Philippine Society for Developmental Biology (PSDB) National Annual Convention.
- 2020.9A review of Takunori Minegishi (about mechanobiology of neuronal migration) was published in Front. Cell Dev. Biol.
- 2020.2Prof. Inagaki gave an invited talk about "forces to drive axon guidance and neuronal migration" at Actin assembly for Intracellular Functions. (See here)
- 2020.2Prof. Inagaki gave an invited talk at International Symposium on AMED "Mechanobiology" Project.
- 2019.8A study of Akihiro Urasaki (about new shootins and shootin-mediated collective cell migration) was published in Scientific Reports.
- 2019.4Prof. Inagaki gave an invited talk at EMBO Workshop Neuronal Cytoskeleton.
- 2019.2A study of Huang Liguo (about Rab33-mediated axon outgrowth) was published in Scientific Reports.
- 2018.10A study of Takunori Minegishi (about the molecular mechanics of shootin1b-mediated neuronal migration) was published in Cell Reports
- 2018.8A study of Kentarou Baba (about the molecular mechanics of netrin-1-induced axon guidance) was published in eLife
- 2018.2A study of Kouki Abe (about a new mechanism of axon guidance) was published in PNAS
- 2017.3A review about actin wave was published in Trends in Cell Biology.
- 2016.12A column including an interview of Professor Inagaki about "actin wave" was peported in PNAS.
- 2016.10Kentaro Baba (D3) won an excellent poster prize in the third "Dynamical ordering of biomolecular systems for creation of integrated functions" young person meeting for the study.
- 2016.5A study of Yasuna Higashiguchi (about a new shootin1, shootin1b) was published in Cell and Tissue Research.
Outline of Research and Education

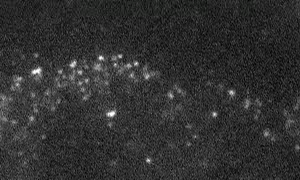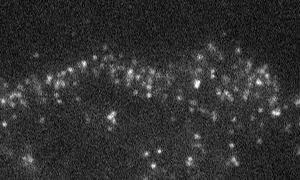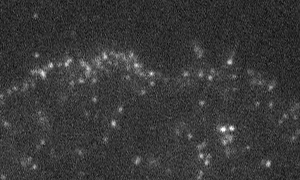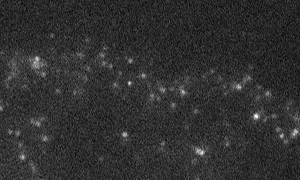
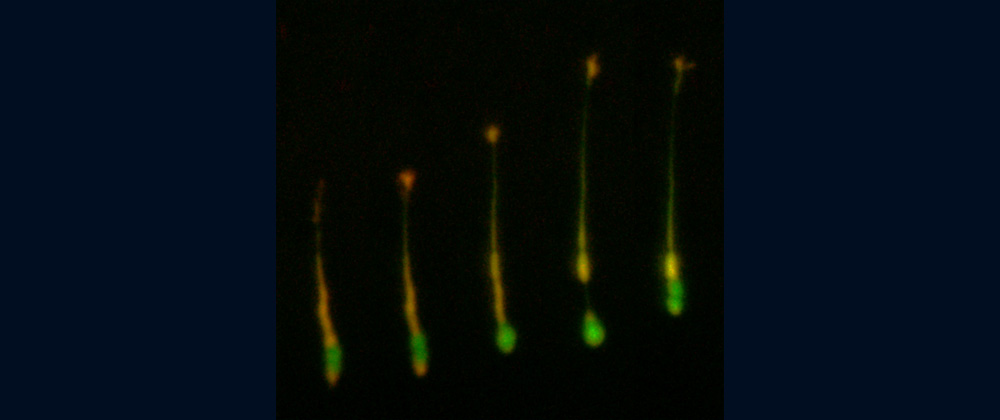
Fig. 1 Shootins move like shooting stars [5,17].
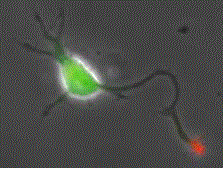
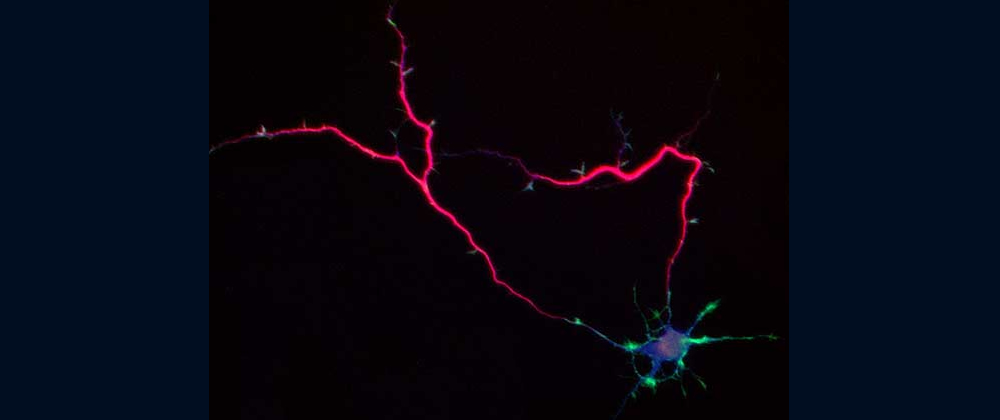
Fig. 2 Shootin1a (red) is a key molecule involved in axon formation and axon guidance [7,8,12,13,17,19].

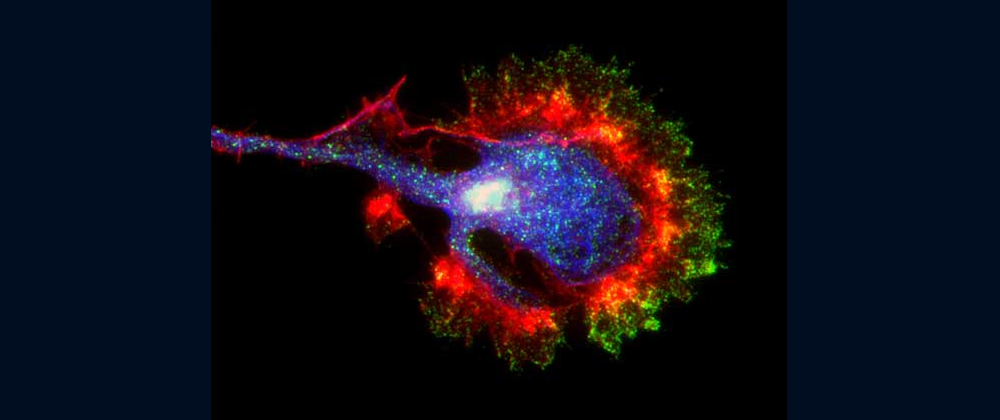
Fig. 3 Shootin1b (magenta) is involved in neuronal migration [6,10].
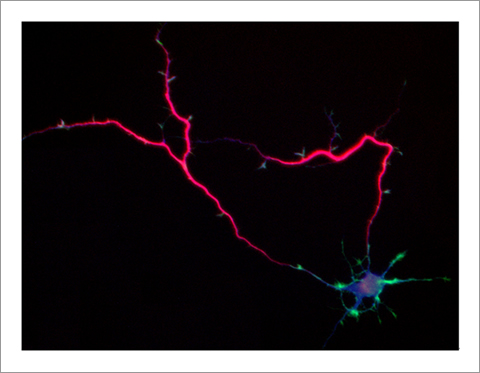
Fig. 4 Singar knockdown leads to formation of surplus axons leading to disruption of neuronal polarity [18].
Fig. 5 Actin wave (red) is a new type of intracellular protein transport system for cell polarization and migration [9,11].
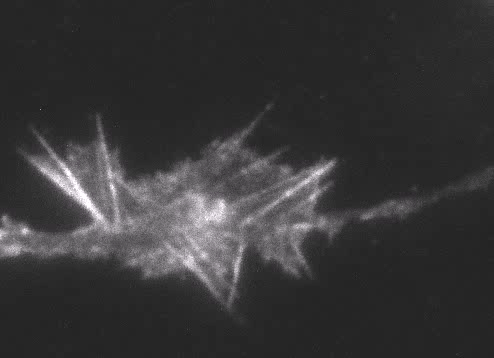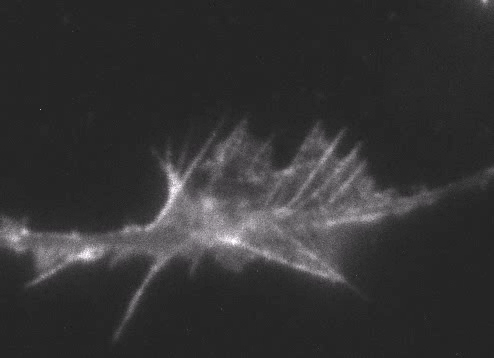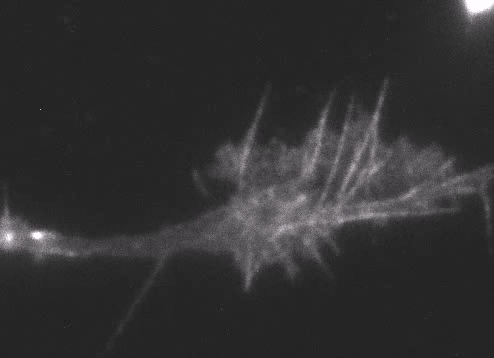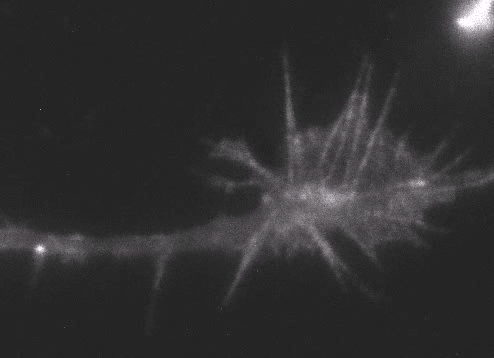
Fig. 6 Actin wave analyzed by single molecule imaging [11].
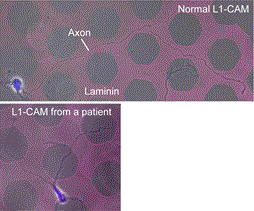
Fig. 7 We recently reported a new type of axon guidance mechanism (Grip & Slip mechanism). This mechanism is disrupted in a human patient of L1-CAM/CRASH syndrome, suffering corpus callosum agenesis and corticospinal tract hypoplasia [8].
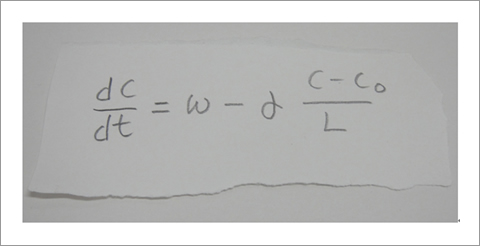
Fig. 8 A mathematical equation to describe shootin1a accumulation in axonal tip [16].

Fig. 9 Shootin1 mediated neuronal polarization (left) is described by a quantitative mathematical model (right) [15.16].
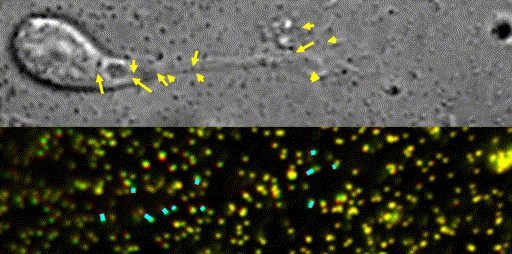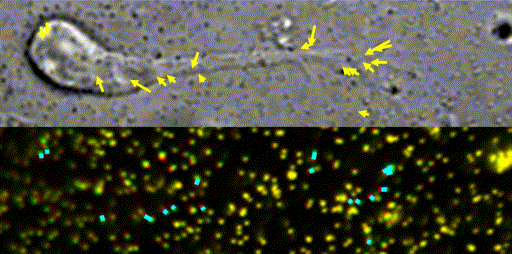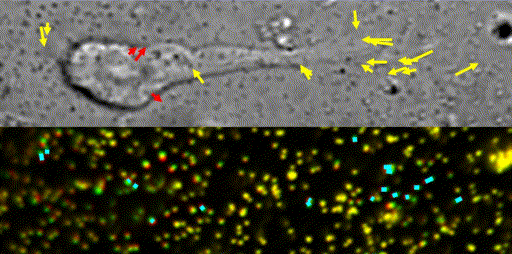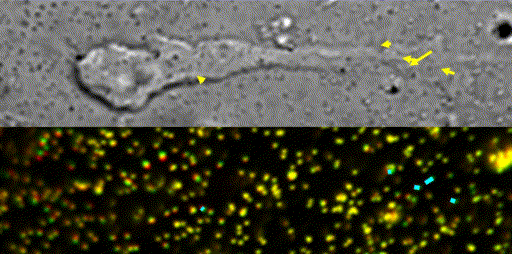
Fig. 10 Traction force microscopy to measure forces for axon guidance [7,8] and neuronal migration [6].
Neurons extend axons, and form elaborate networks in our brain; all the brain activities depend on neuronal networks. To establish proper neuronal networks, axons decide their migratory route in response to gradients of chemical signals in the brain with extraordinary sensitivity. But, little is known about how axons are able to decide their migratory routes by reading gradients of chemical signals and by translating them into directional driving force.
In addition to axons, various cells migrate within our body, thereby playing key roles in organ formation, immune responses, wound healing and regeneration. Disruption of axon guidance and cell migration is implicated in diseases, including birth abnormality, neuronal disabilities, immune disorders and cancer metastasis.
Our laboratory focuses on the proteins, Shootin1a (Fig. 1 and 2) [12,21], Shootin1b (Fig. 2) [8] and Singar (Fig. 4) [20], which we identified by proteome analyses [22-24], as well as their interacting proteins, Cortactin [14], L1-CAM [10] and Rab33 [6,16]. We are analyzing the molecular mechanisms for axon formation [16-21,25,26], axon guidance [9,10], cell migration [8], and synaptic plasticity [27]. We also analyze actin waves which is a new type of intracellular protein transport system for cell morphogenesis and cell migration (Fig. 5 and 6) [11,13].
We expect that our studies will help us to understand the mechanisms underlying neuronal morphogenesis and neuronal network formation, as well as the mechanisms underlying diseases including birth abnormality [10] (Fig. 7), neuronal disabilities, neuropsychiatric disorders and immune disorders, giving us a new window into therapeutic strategies for nerve injury, Alzheimer’s disease, neuropsychiatric disorders and cancer metastasis.
In our laboratory, we use standard biological methods (biochemistry, molecular biology, cell biology and developmental biology), as well as cutting-edge technologies including single molecule imaging [8-10,14,15,19], systems biology (Fig. 8 and 9) [13,17,18], mechanobiology (Fig. 10) [8-10,13,14,18], and laser manipulation [13,19]. Yet, we do not ask previous experiences to join our laboratory. We will educate these methods from basics step-by-step: starting research in our laboratory is not difficult.
Major Research Topics
1) Axon/dendrite formation, neuronal polarization
The basic function of neurons is to receive signals, integrate them and transmit them to the next neuron. Thus, neurons can function as semiconductors in the brain. Neuronal polarity plays an essential role for this function. Namely, most of neurons form a single axon and multiple dendrites. Dendrites receive signals from outside, while an axon output signals. Then a signal flow arises from dendrites via cell body to the axon. How neurons acquire neuronal polarity? Our laboratory investigates the mechanism for axon/dendrite formation and neuronal polarization, focusing on Shootin1a (Fig. 2) [18,21], Singar (Fig. 4) [20] and Rab33 [6,16].
2) Mechanical forces for axon guidance and cell migration
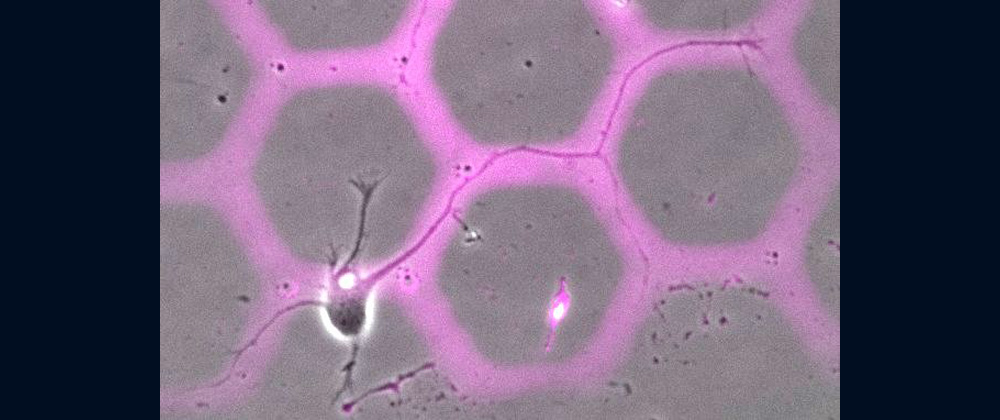
Animals move and change their shape, using force generated by muscles. How cells do so? Actin polymerizes near the leading edges of neurite tips and migrating cells. We reported that shootin1a interacts with both the polymerizing actin filaments and the cell adhesion molecule L1-CAM at neurite tips, thereby generating traction force for neurite outgrowth [19]. We are currently analyzing molecular mechanisms for generation and regulation of mechanical forces (Fig. 10) for axon guidance [9,10] and neuronal migration [5,8]. We also reported a new type of axon guidance mechanism (Grip & Slip mechanism) regulated by the extracellular matrix protein Laminin [10] and substrate stiffness [3].
3) Molecular mechanism for synaptic plasticity
Synaptic plasticity is a key process underlying learning and memory. We are analyzing the molecular mechanics for synaptic plasticity, focusing on the cell signaling and morphological change of dendritic spines [4, 27].
4) Actin waves: a new mechanism for cellular protein transport
Actin filaments and associated proteins undergo wave-like movement in various cell types (Fig. 5 and 6) [11]. We reported that axonal actin waves migrate by means of directional assembly and disassembly of membrane-anchored actin filaments [13]. Thus, the actin wave in axons represents a new type of machinery that translocates actin and associated proteins to the cell edge. The involvement of actin waves in axon outgrowth was also demonstrated by laser-manipulation of the adhesion substrate [13]. We are currently analyzing roles of actin waves in cell polarity formation and migration, introducing single molecule imaging and mathematical modeling (Fig. 8) [11,17,18].
5) Research in medicine: brain diseases and cancer metastasis
We expect that our studies will help us to understand the mechanisms underlying diseases including birth abnormality, neuropsychiatric disorders, and immune disorders, giving us a new window into therapeutic strategies for nerve injury, Alzheimer’s disease, neuropsychiatric disorders, and cancer metastasis. We recently reported “Grip & Slip mechanism” for axon guidance; this mechanism is disrupted in a human patient of L1-CAM/CRASH syndrome, suffering corpus callosum agenesis and corticospinal tract hypoplasia [10]. We are also currently analyzing the molecular mechanisms for immune cell migration and tumor cell metastasis.
References
[1] Minegishi T. et al., J. Vis. Exp., e63227, 2021 (LINK)
[2] Kastian R. et al., Star Protoc., 2, 100904, 2021 (LINK)
[3] Abe K. et al., Biophys. J., 120, 3566-3576, 2021 (LINK)
[4] *Kastian R. et al., Cell Rep., 35, 109130, 2021 (LINK)
[5] Minegishi T. et al., Front. Cell Dev. Biol., 8, 863, 2020 (LINK)
[6] *Huang L. et al., Sci. Rep., 9, 1799, 2019 (LINK)
[7] Urasaki A. et al., Sci. Rep., 9, 12156, 2019 (LINK)
[8] *Minegishi T. et al., Cell Rep., 25, 624-639, 2018 (LINK)
[9] *Baba K. et al., eLife, 7, e34593, 2018 (LINK)
[10] *Abe K. et al., PNAS, 115, 2764-2769, 2018 (LINK)
[11] Inagaki N. and Katsuno H., Trends Cell Biol., 27, 515-526, 2017 (LINK)
[12] *Higashiguchi et al., Cell Tissue Res., 366, 75-879, 2016 (LINK)
[13] *Katsuno H. et al., Cell Rep., 12, 648-660, 2015 (LINK)
[14] *Kubo Y. et al., J. Cell Biol., 210, 663-676, 2015 (LINK)
[15] Toriyama M. et al., Curr. Biol., 23,529-534, 2013 (LINK)
[16] *Nakazawa H. et al., J. Neurosci., 32, 12712-12725, 2012 (LINK)
[17] Inagaki N. et al., Dev. Neurobiol., 71, 584-593, 2011 (LINK)
[18] Toriyama M. et al., Mol. Syst. Biol., 6, 394, 2010 (LINK)
[19] Shimada T. et al., J. Cell Biol., 181, 817-829, 2008 (LINK)
[20] *Mori T. et al., J. Biol. Chem., 282, 19884-19893, 2007 (LINK)
[21] *Toriyama M. et al., J. Cell Biol., 175, 147-157, 2006 (LINK)
[22] Inagaki and Katsuta, Curr. Proteomics 1, 35-39, 2004
[23] **Nomura E. et al., J. Mass Spectrometry, 39, 666-672, 2004
[24] **Oguri T. et al., Proteomics, 2, 666-672, 2002
[25] Fukata Y. et al., Nature Cell Biol., 4, 583-591, 2002
[26] Inagaki N. et al., Nature Neurosci., 4, 872-873, 2001
[27] Inagaki N. et al., J. Biol. Chem., 275, 27165-27171, 2000 (LINK)
(*Papers by doctor course students; **papers by master course students)
Contact us
Naoyuki Inagaki, MD, PhD
Graduate School of Biological Sciences
Nara Institute of Science and Technology
Ikoma 630-0192, Japan (access map)
Tel: +81-743-72-5466
Email: ninagaki(a)bs.naist.jp

{kind=link}
{kind=link}
{kind=link}