幹細胞の増殖・分化・老化の制御機構
花幹細胞制御
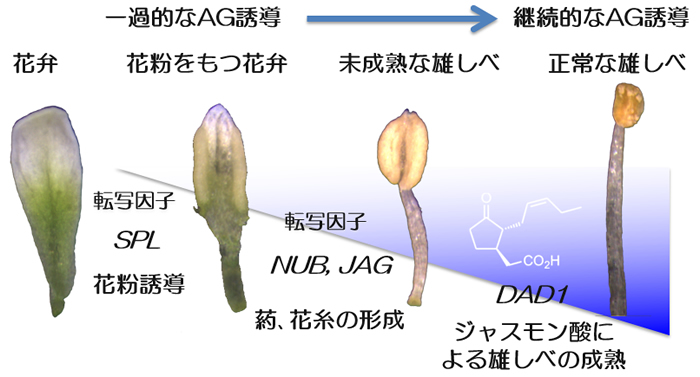
花のホメオティックタンパク質AGAMOUSは、花幹細胞の増殖抑制を行うと同時に、雄しべや雌しべなどの生殖器官の分化誘導を行います。
私たちは、これまでに、花器官の分化において、AGAMOUSが生殖器官の発生過程に継続的に発現して、さまざまな下流遺伝子の制御をしていることを明らかにしてきました。
(Ito et al. Nature 430:356-360, 2004; Ito et al. Plant Cell 19:3516-3529, 2007)

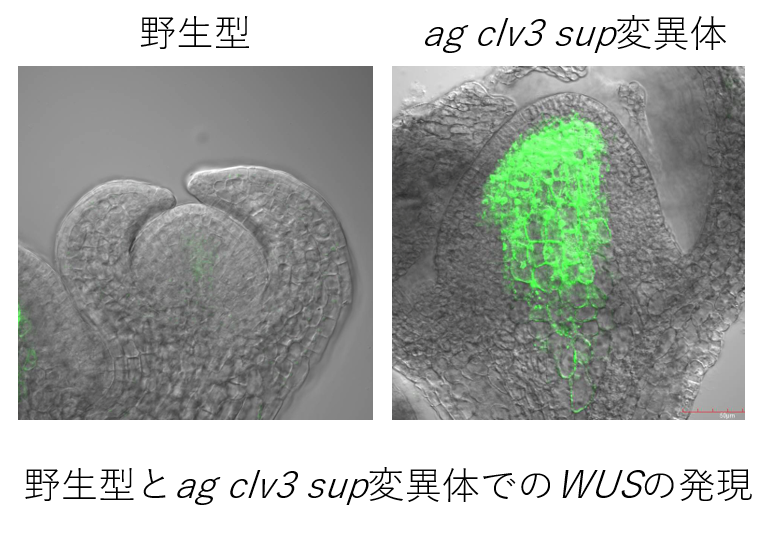
これまでに、花の幹細胞活性の抑制にかかわる複数の遺伝子が知られていますが、それらの分子作用メカニズムはまだまだわかっていません。花幹細胞の増殖抑制と分化制御にかかわる転写制御因子であるSUP やCRC の下流の解析を行っています。それらのターゲット遺伝子を同定し、時空間特異的な発現制御機構とターゲット因子の作用機構の研究を行うことにより、花幹細胞の増殖抑制経路の制御機構を解明していきます。
マスター制御因子による2つの異なる細胞の運命決定機構
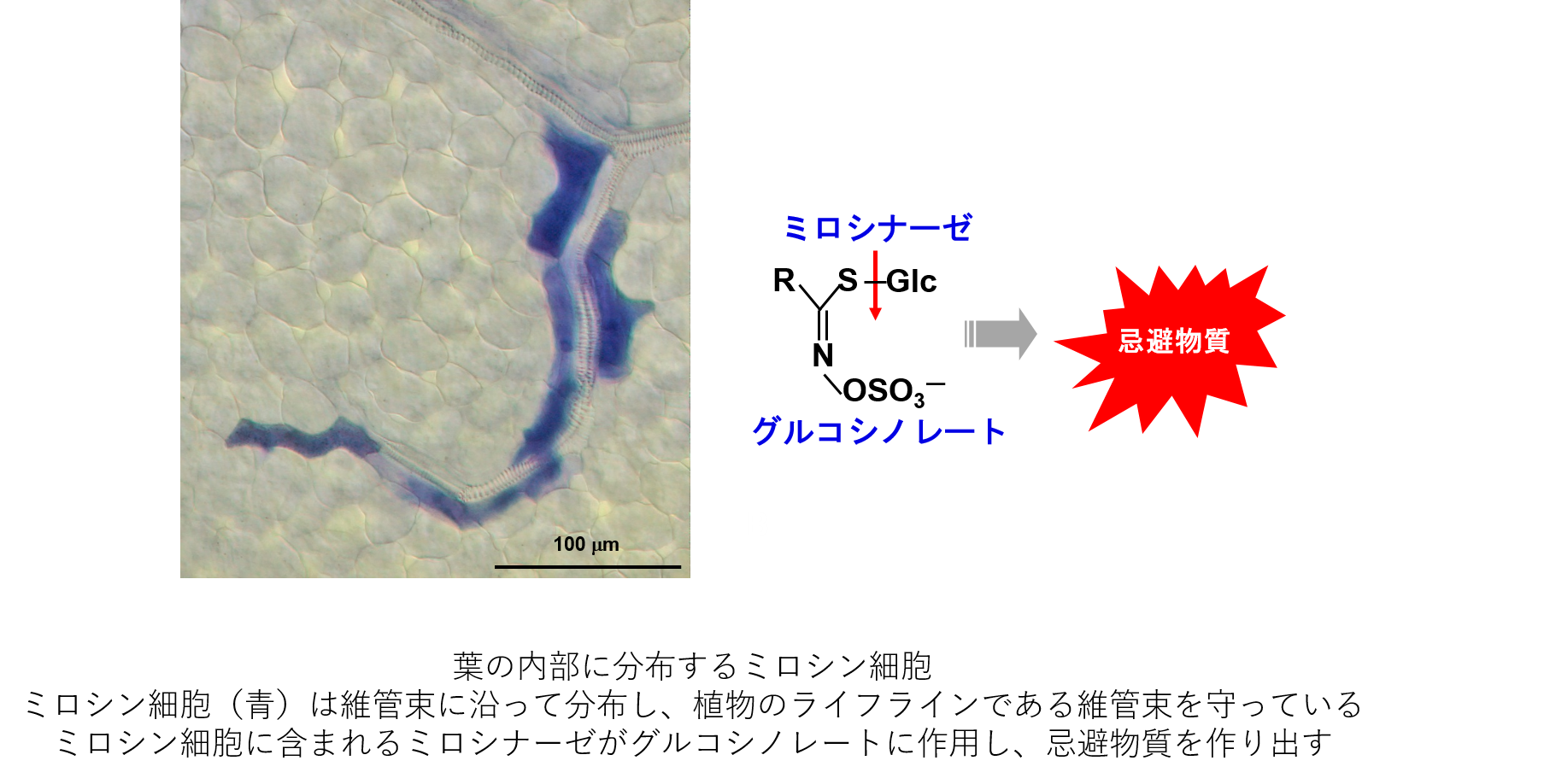
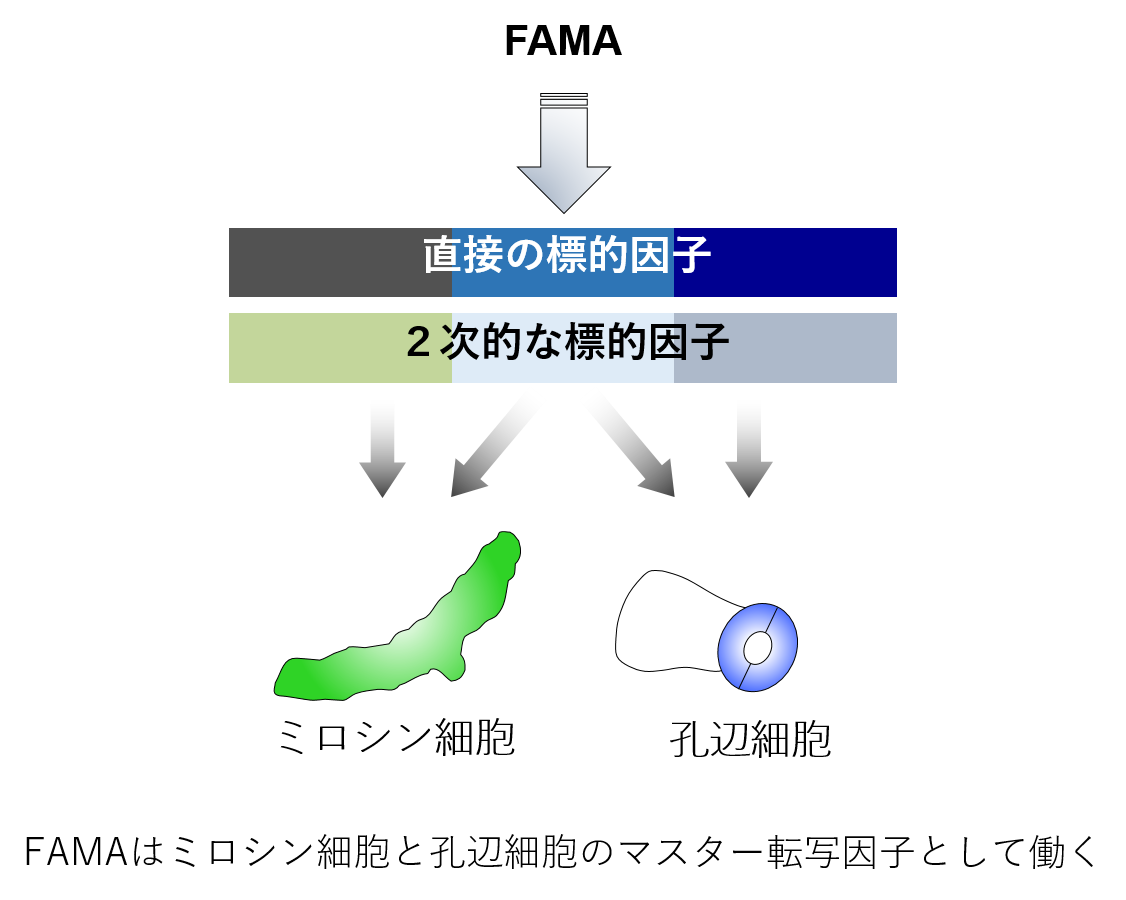
細胞の分化は、主としてマスター転写因子によって制御されています。bHLH型転写因子FAMAは、二酸化炭素の取り込みを担う孔辺細胞分化のマスター制御因子であるのに加えて、生体防御に働く異形ミロシン細胞のマスター制御因子でもあります。孔辺細胞は植物の表面に、ミロシン細胞は植物の内部に形成されます。しかし、FAMAがどのように2つの細胞を作り分けているのか詳細なメカニズムは不明です。FAMA遺伝子座のエピジェネティックな制御機構とFAMAの相互作用因子に着目することで、2つの異なる細胞運命の分岐点を明らかにしたいと考えています。加えて、順遺伝学的手法によりミロシン細胞分化の新規制御因子を探索しています。