
2024.10.03
植物ホルモンによる苔類の器官形成制御メカニズム
植物成長制御研究室・助教・安喜史織
- 要旨
- 植物ホルモンのひとつであるサイトカイニンは植物の成長に重要であることが古くから知られていますが、サイトカイニンシグナル下流の分子メカニズムについてはまだまだ不明な点が多く残っています。私たちはこれまで苔類ゼニゴケをモデルに、サイトカイニンによる植物の形づくりのしくみを理解するべく、研究を進めてきました。本稿では、ゼニゴケにおけるサイトカイニンの役割とその分子メカニズムについて紹介します。
- 主要関連論文
- Aki SS, Morimoto T, Ohnishi T, Oda A, Kato H, Ishizaki K, Nishihama R, Kohchi T, Umeda M. R2R3-MYB transcription factor GEMMA CUP-ASSOCIATED MYB1 mediates the cytokinin signal to achieve proper organ development in Marchantia polymorpha. Sci Rep. 2022, 12(1):21123.
doi: 10.1038/s41598-022-25684-3.
1.はじめに
植物は周辺の環境に応答しながら根や茎を成長させて葉を作り、ときに花を咲かせます。このような形づくりには植物ホルモンの働きが重要です。サイトカイニンは細胞分裂、器官形成、老化など幅広い生命現象を制御する植物ホルモンです。サイトカイニンシグナル伝達はヒスチジンキナーゼであるサイトカイニン受容体により開始され、そのシグナルがリン酸リレーを介して転写因子型レスポンスレギュレーター(type-B RR)へと伝達されて、さまざまな遺伝子の転写が活性化されます。Type-B RRの標的遺伝子のひとつである非転写因子型レスポンスレギュレーター(type-A RR)はサイトカイニンシグナルを抑制することが知られています。これまでの知見の多くは被子植物を用いた研究により明らかにされたことですが、実は被子植物の多くは非常に遺伝子の冗長性が高く、サイトカイニンシグナル伝達を構成する遺伝子を数多く持っています。そのため、サイトカイニンシグナルの下流でどのような因子が個々の現象を制御しているのかを明らかにすることは困難でした。例えば、サイトカイニンによる細胞増殖の制御においては、サイクリンD3(CYCD3)が主要な役割を果たすと考えられていますが、どのtype-B RRがCYCD3の発現を誘導するのか、またCYCD3が細胞周期におけるサイトカイニンの作用にどの程度寄与するのか、など多くの疑問が残されています。
2.苔類ゼニゴケを用いたサイトカイニンシグナルの解析
苔類ゼニゴケは陸上植物の基部に位置するという点で近年注目されているモデル植物です。2017年には全ゲノム配列情報が決定され、陸上植物のもつ転写因子やシグナル伝達因子などをコードする制御系の遺伝子をほぼ一通りもつこと、またそれらの冗長性が比較的低いことが明らかになりました1)。例えば、シロイヌナズナはtype-B RRおよびtype-A RRをコードする遺伝子をそれぞれ10以上持っていますが、ゼニゴケは1つずつ(MpRRB、MpRRA)しか持っていません 1), 2)。そこで、MpRRBやMpRRAのノックアウト株、過剰発現株、内生サイトカイニン量を低下させた株などを作成して解析したところ、サイトカイニンが無性生殖器官である杯状体の形成を促進する一方で、個体の支持を担う仮根の形成を阻害するなど、ゼニゴケの器官形成を制御することが明らかになりました2), 3)。ゼニゴケは葉状体先端のくぼみ(ノッチ)に幹細胞を1つ持っており、個体を構成するすべての細胞は幹細胞から生まれる4つの細胞系譜(背側、腹側、両側方)に由来します。MpRRBはノッチ付近で蓄積しており、また内生サイトカイニン量を低下させた株のノッチ付近を詳細に観察したところ、幹細胞から背側方向に数細胞進んだところで起こる杯状体原基の形成が行われていないことが明らかになりました。このことから、サイトカイニンはゼニゴケにおいて幹細胞由来の前駆細胞の増殖あるいは分化を制御することで、杯状体や仮根の形成に重要な役割を担っていることが示唆されました(図1)。
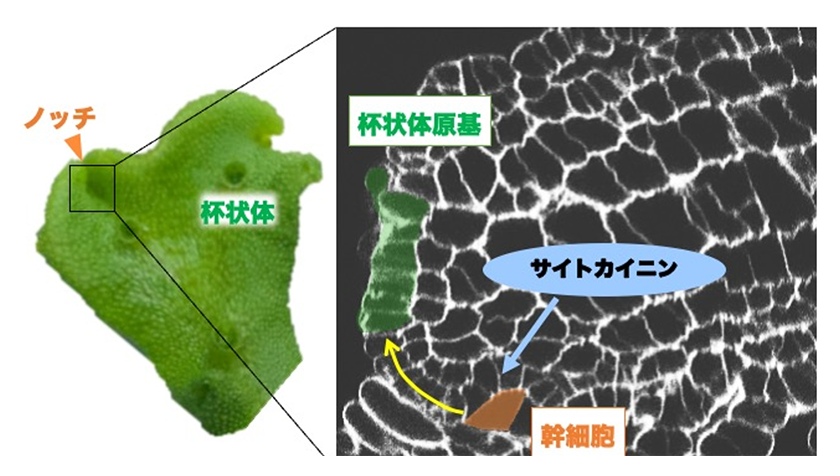
図1.サイトカイニンシグナルによる杯状体の形成促進メカニズム
3.サイトカイニンシグナルと杯状体形成をつなぐ因子
さらにサイトカイニンがどの様に器官形成を制御するのか、その分子メカニズムを明らかにするため、サイトカイニンによる杯状体形成の促進に着目し、すでに杯状体形成の鍵因子であることが報告されていたR2R3型MYB転写因子GEMMA CUP-ASSOCIATED MYB1(MpGCAM1)との関係について調べました。その結果、MpGCAM1の発現がサイトカイニンやサイトカイニンシグナルの増加に応答して上昇することが明らかになりました。さらに、MpGCAM1とMpRRAの二重変異株では、MpRRAの欠損によりサイトカイニンシグナルが上昇しているにも関わらず、杯状体が形成されませんでした。このことから、MpGCAM1はサイトカイニンシグナルの下流で機能することが遺伝学的に明らかになりました。また、MpGCAM1の誘導型過剰発現株は未分化な細胞塊を形成することが報告されていましたが、これは野生株、MpRRBのノックアウト株両方の背景で観察されました。しかしながら、MpGCAM1の誘導を止めた後に細胞塊から形成される葉状体は、野生株よりもMpRRBのノックアウト株背景の方が小さいことがわかりました。葉状体は、頂端ノッチが新たに形成された後、ノッチ付近の細胞が増殖することで成長します。今後さらなる解析によりサイトカイニンシグナルがどのプロセスに関与するのか明らかにしたいと考えています。
4.おわりに
サイトカイニンシグナルの下流で機能するMpGCAM1のオルソログは被子植物にも保存されており(シロイヌナズナではRAX)、RAXは腋芽の形成に重要であることが報告されています。しかし、ゼニゴケではサイトカイニンがMpGCAM1を制御するのに対して、RAXはサイトカイニンの生合成を促進します。すなわち、RAXはサイトカイニンの上流で機能することから、進化の過程で両者の関係性が逆転したと考えられます(図2)。MpGCAM1をシロイヌナズナのrax変異体で発現すると腋芽形成の欠損が部分的に回復することを考慮すると、RAXオルソログは進化の過程で標的遺伝子のプロモーター配列を変更したのかもしれません。また最近、光情報に応答して形成される有性生殖器官においてもサイトカイニンシグナルが重要な役割をもつことがわかってきました5)。今後は、サイトカイニンシグナルの下流因子の解析にとどまらず、ノッチ付近の細胞のライブイメージングやシングルセルRNAシーケンスなどを駆使して周辺環境に応答した形づくりへの関与や他のホルモンとのクロストークなど、植物が進化の過程で獲得したしなやかな生存戦略について理解を深めていきたいと思っています。
本研究を進めるにあたり,所属研究室の梅田正明教授、河内孝之教授(京都大学)、石崎公庸教授(神戸大学)、西浜竜一教授(東京理科大学)、榊原均教授(名古屋大学)、経塚淳子教授(東北大学)、楢本悟史准教授(北海道大学)をはじめ多くの方々にお世話になりました。この場をお借りし厚く御礼申し上げます。
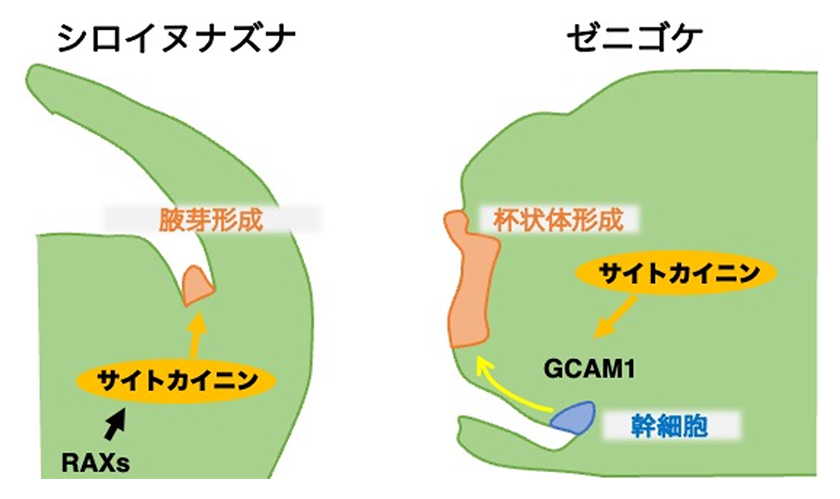
図2.シロイヌナズナとゼニゴケにおけるR2R3型MYB転写因子とサイトカイニンの関係性
参考文献

著者
安喜 史織 Researchmap
略歴
2011年京都大学大学院理学研究科博士後期課程修了。企業を経て2013年から奈良先端科学技術大学院大学博士研究員、2020年4月より現職
- 研究内容:植物の器官形成メカニズム
- 抱負:人生を楽しむ
- 関心ごと:個人の可能性を最大限に引き出す方法
安喜 史織 NAIST Edge BIO, 0027. (2024)