
2024.05.01
ミヤコグサが種内に持つ表現型や遺伝子型の多型と適応進化機構の解明
バイオエンジニアリング研究室・助教・若林智美
- 要旨
- 生物多様性は、小さな進化の積み重ねで形成されてきています。本研究で対象としているミヤコグサは、日本では沖縄県宮古島から北海道利尻島まで広く連続的に分布しているマメ科の草本植物です。日本は南北に長い地形や山脈を多く持つことなどの様々な要因により、環境勾配の大きな地域です。様々な地域に由来する系統を観察すると、本種の種内には、開花時期や茎の色、葉の大きさや形など、様々な形質に違いが見られます。私たちは、その中でも特に繁殖成功に関わるいくつかの形質に着目して、適応進化の仕組みを明らかにしようと試みています。本種はマメ科のモデル植物の一つで、全ゲノムデータや、各地から集められた系統とその遺伝子型情報などを、野生生物の適応進化研究に利用できる貴重な材料です。私たちは、これらの研究資源を活かして、関連する環境要因の推定や遺伝的要因の検出、さらには自然選択の影響の推定により、包括的にミヤコグサの適応進化のプロセスの解明に挑戦しています。本稿では特に、次の世代の種子生産に直結する開花時期の違いに着目した研究について紹介します。
- 主要関連論文
- Shah N, Wakabayashi T, Kawamura Y, et al. (2020) Extreme genetic signatures of local adaptation during Lotus japonicus colonization of Japan. Nature communications. 11, 253.
doi: 10.1038/s41467-019-14213-y - 若林 智美, 瀬戸口 浩彰. (2019) ミヤコグサ国内野生系統が示す開花時期の違いとその遺伝基盤の解明 〜全ゲノム関連解析による責任遺伝子の検出〜. 植物科学最前線 10: 190-195.
doi: 10.24480/bsj-review.10c10.00170
1.生物の多様性と適応進化
地球上には様々な気候条件が存在し、多様な生物が生息または生育しています。生物種や、生物個体同士は相互に関わり合い、複雑な生態系が形成されてきています。特に、一度根を張るとその場から動くことのできない植物にとって、生育地環境は個体の生存や繁殖成功に大きく関わる重要な要素です。1993年に発効された生物多様性条約では、生物多様性を「生態系の多様性」、「種の多様性」、「遺伝子の多様性」の3つの区分で定義しています。この中でも「遺伝子の多様性」は、種内に存在する遺伝的な多様性のことを指しており、様々な生物種において個体群が地理的または生殖的に隔離されることや、環境適応などと関連して形成されます。
適応進化は生物が生息または生育する環境に応じて、世代を経て性質が変化する現象のことを言います。気候変動や、生物が分布を変化させていく中で、異なる環境に置かれた生物集団では絶えずこの適応進化が起こっています。例えば、日本列島のような様々な環境が存在する地域に広く分布する生物種では、それぞれの地域環境に応じた性質を獲得することや、地理的に遺伝的な交流が妨げられることから、同じ種であっても種内に多様性が生まれます。
2.ミヤコグサの過去の分布変遷と種内の多様性
私たちが研究対象としているミヤコグサ(Lotus japonicus)は、春になると黄色い花をつけるマメ科の草本植物で、道端や草地などでも見つけることができる身近な植物です。国内だけでも北は北海道利尻島から南は沖縄県宮古島まで広く連続的に分布しています。日本列島は南北に広い地形を有しており、山脈が多いことや海に囲まれていること、季節風の影響を受けるなどの様々な要因により、環境勾配の大きな地域です。様々な環境が存在する日本列島に広く分布しているミヤコグサは、各地の自生地環境に適した表現型を獲得するような適応進化をしてきていると考えられます。
ミヤコグサは種内に、開花時期や茎の色、地上部の形態など、様々な形質の種内多型を持つことが知られています(Hashiguchi et al., 2011など)。実際に国内各地の自生地から集められたミヤコグサの野生系統を栽培し、観察してみると、種内に様々な形質の表現型多型を持つことがわかります。例えば、播種してから最初の花が咲くまでの日数には顕著な違いが見られます(図1)。ミヤコグサは長日植物で、ある一定の長さよりも長い日長を認識して花成が誘導されます。南北に長い日本列島では、大まかに言えば南北で日長に違いがあり、それに付随してミヤコグサ種内に見られる開花所要日数の違いも、由来地の緯度に沿って南由来の系統から順に早咲きとなります。また、目に見える形態などの多様性に加え、ゲノム全体に塩基配列の違いもあることがわかっています。私たちはこれらの多型情報を利用して、ミヤコグサが日本列島各地の環境に適応してきたプロセスを明らかにできないかと考えました。
そこで私はまず、国内外の研究者たちと連携して、ミヤコグサの日本列島における過去の分布変遷を推定しました。国内の分布を網羅するような野生系統を用いて、そのゲノム網羅的な遺伝子型多型に基づいた集団遺伝学的解析を行いました。その結果、日本のミヤコグサは約1万年前の最終氷期後の気候変動に伴い、対馬付近から九州を経由して、大きく分けて3つの経路で日本列島に広がったことが示されました(図2、Shah et al., 2020)。このような足取りで様々な環境に移入を繰り返しながら日本列島全体に分布を広げたミヤコグサを対象に、私たちは特に繁殖成功に関わる形質の表現型多型に着目した研究を展開してきています。
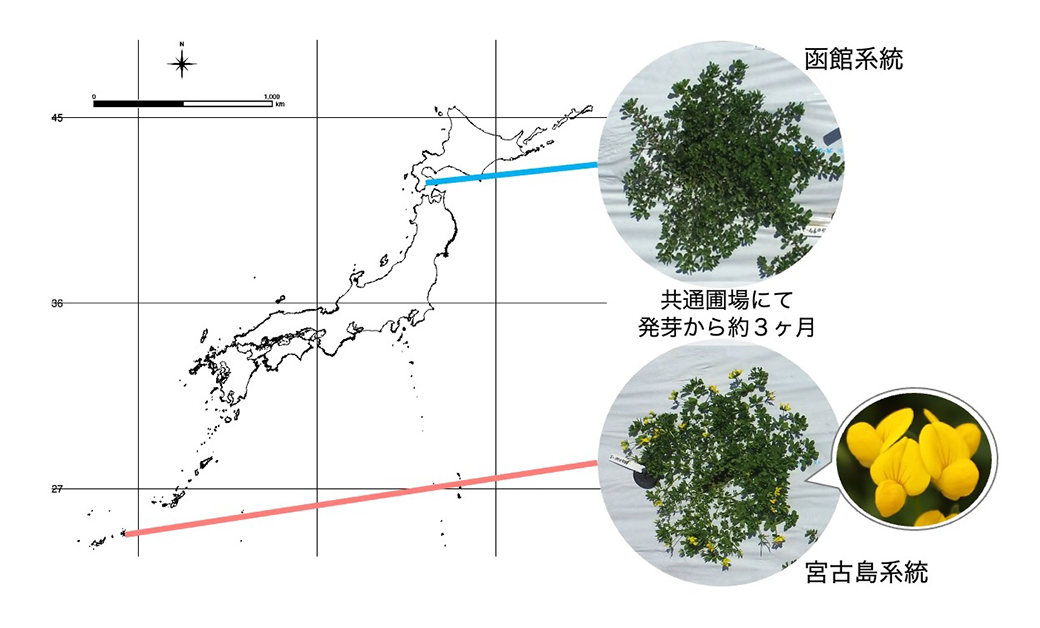
図1.ミヤコグサの種内に存在する開花時期多型
同じ条件で栽培し、同じ日に観察した函館系統と宮古島系統を例に比較すると、宮古島系統では花がたくさんついている一方で、函館系統ではまだ栄養成長の段階である様子が観察できます。
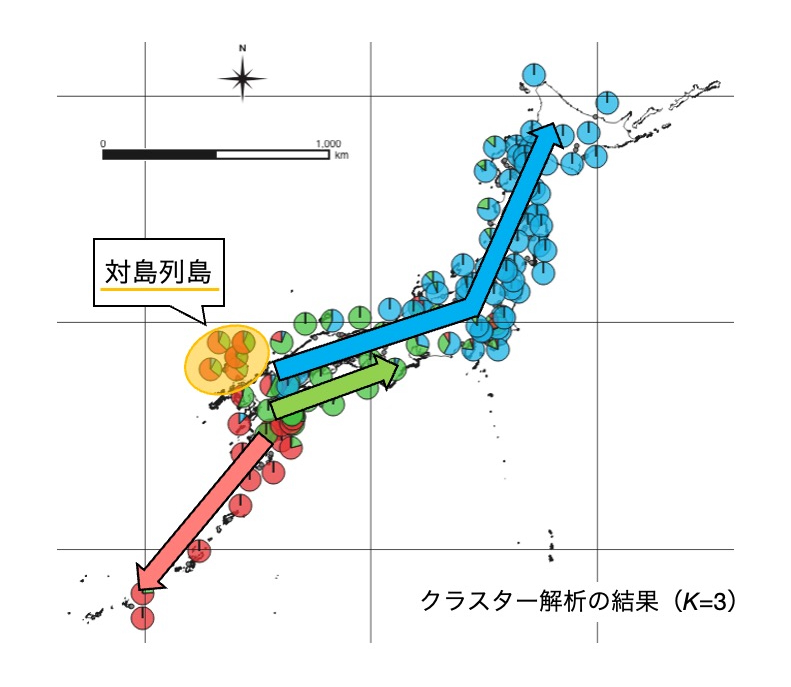
図2.日本列島におけるミヤコグサの過去の分布変遷
日本に分布するミヤコグサは、最終氷期後の気候変動に伴い大きく分けて3つの経路で日本列島に分布を広げたと考えられます。
3.開花時期の多様性と環境適応
本稿では、特に次代の繁殖成功に直結する開花時期の多型に焦点を当てた研究について説明します。先に述べたように、ミヤコグサは種内に開花時期の多型を持ちます。この表現型多型について、まず、関連する気候要素や遺伝的要因を明らかにすることを目標に、次の実験と解析を試みました。栽培実験では、日本各地に由来する100系統以上の野生系統を同一条件で栽培し、開花までの日数を計測しました。開花所要日数データは、共同研究者による異なる条件下でのデータと合わせて解析することで、ミヤコグサの開花時期の違いには、主に栄養成長期間と日長が関わっていることが示唆されました。また、全ゲノム関連解析により関連する遺伝領域の検出を行い、複数の候補遺伝子を挙げてきています。この中の特に有力なものについては、ゲノム編集により遺伝子を不活化して表現型への効果を実験的に示しました。これら複数の遺伝子の遺伝子型の違いにより、本種の連続的な開花時期多型がもたらされている可能性が考えられます(図3)。
さらに、候補遺伝子の遺伝子領域にかかる自然選択の影響についても、ゲノム網羅的な塩基多型情報を用いて解析しました。その結果、候補遺伝子のうちの一つについては、その遺伝子領域のうち非コード領域の塩基配列に平衡選択という自然選択が強く影響してきたことがわかりました。平衡選択は、複数の遺伝子型を種内もしくは集団内に維持する方向に働く自然選択です。その一方で、コード領域上の変異が排除されるような浄化選択が働いたことも示す結果となっており、コードされるアミノ酸配列の違いではなく、遺伝子の発現量やスプライシングバリアントなどによる表現型への影響が考えられました。今後は、この遺伝子の野生系統に共有されている遺伝子型間で、遺伝子発現量やスプライシングバリアントの有無などを解析し、多型が維持されるより詳細な仕組みを明らかにしていきたいと考えています。
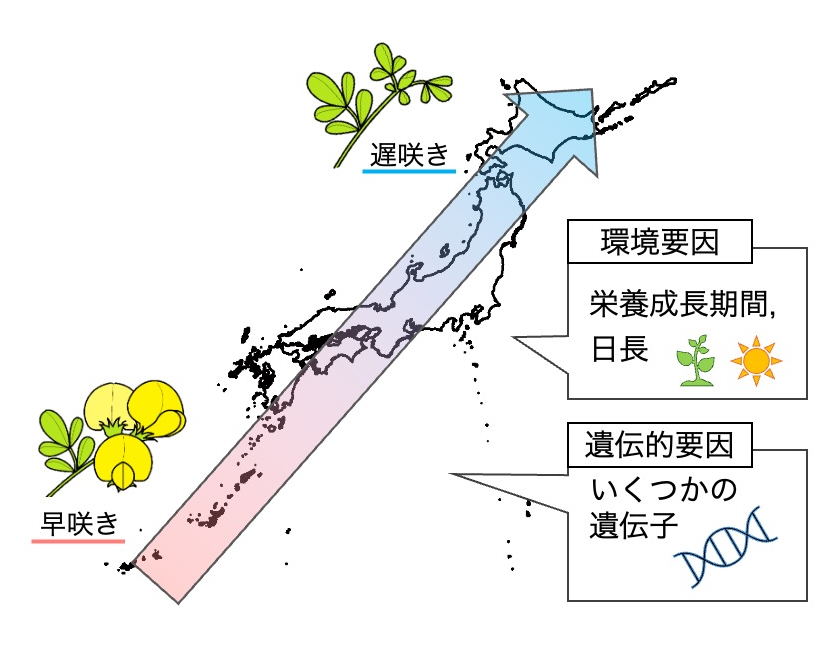
図3.ミヤコグサの開花時期多型に関わる要因
ミヤコグサの開花時期には、栄養成長期間と日長が環境要因として関わること、さらには検出されたものを含むいくつかの遺伝的要因の組み合わせによって、連続的な表現型多型が形成されている可能性が示されました。
4.植物の環境適応の仕組みを包括的に理解するために
本稿では述べていませんが、最近では、開花時期以外にも地上部バイオマスに関わる形質や、一年生と多年生の生活型の違いなどにも着目して研究を展開しています。地上部バイオマスに関わるような複数の形質の表現型を解析することで、ミヤコグサの地上部戦略のパターンが複数あることが明らかになり、表現型多型に関わる遺伝的要因を検出することができています。一年生と多年生の違いについても、関連する環境条件を推定し、遺伝領域の検出も行ってきました。開花時期を含め、これらの形質は互いに関連し合っています。今後はそれぞれの形質についての研究を遺伝子発現のレベルでも解析するほか、形質ごとに得られた結果を統合することで、ミヤコグサが日本列島全体に分布を拡大しながら環境適応し、種内に多様性を形成してきた仕組みを包括的に明らかにすることを目指したいと考えています。
また、紹介したこれらの研究を進めるにあたり、加藤晃教授(奈良先端大)をはじめ、瀬戸口浩彰教授(京都大学)、川口正代司教授(基礎生物学研究所)、佐藤修正教授(東北大学)、Stig U. ANDERESEN准教授(Aarhus大学)、壽崎拓哉准教授(筑波大学)などの多くの方々に大変お世話になりました。この場をお借りして厚く御礼申し上げます。
参考文献

著者
若林 智美 Researchmap
略歴
2020年京都大学大学院人間・環境学研究科博士課程修了、奈良女子大学理系女性教育開発共同機構 特任助教を経て、2021年11月より現職
- 研究内容:
ミヤコグサの種内多型と多様性の形成や適応進化についての研究に従事。産地の異なるミヤコグサの様々な違いを観察することを日々楽しんでいます。
若林 智美 NAIST Edge BIO, 0024. (2024)