研究プロジェクト
1. 植物がもつ組織とゲノム恒常性の維持機構
樹木が数千年生き続けるように、植物は永続的な生命力をもっています。また、生き続ける中で新たな器官を作り続けます。動物では多能性幹細胞が受精後間もなく消滅してしまいますが、植物ではそれがずっと維持され、増えていきます。そのため、新たな器官を一生作り続けることができるのです。しかし、植物はがんを作りません。なぜ器官発生の過程で過剰な細胞増殖やDNA変異が生じないのか、といった疑問は、未だに未解決の問題として残っています。
私達はこの疑問に答えるために、過剰な細胞増殖を人工的に誘発する実験系を構築しました。これを使って時系列を追って遺伝子発現やホルモンの動きを調べれば、植物が過剰な細胞増殖を抑え込む仕組みを理解できると考えています。また、DNA変異の発生を防ぐには、細胞分裂の過程で生じるDNA複製ストレスを抑え込む制御系が重要だろうと考えています。これもホルモンによって制御されていることが明らかになりつつあるので、「植物ではホルモンが器官発生とゲノム恒常性を協調的に制御することでがんの発生を防いでいる」という仮説を立てています。これを証明することで、植物がもつ永続的な生命力を理解しようとしています。
この研究は、科学研究費補助金・国際先導研究「植物の自律的な器官形成を支える情報分子ネットワークの解明」において、1細胞解析などの最先端技術や理論生物学も取り入れながら進めています。
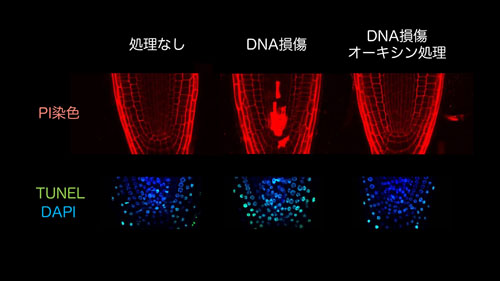
図1. シロイヌナズナの根にDNA損傷を与えると、幹細胞特異的な細胞死が起こる(上段の赤く染まった部分)。しかし、オーキシン処理を施すと、DNA損傷による幹細胞死が完全に抑制される。これらの結果から、オーキシンは植物のゲノム恒常性維持に関与していることが明らかになりつつある。
2. 環境ストレスに応答した細胞分裂の抑制機構
植物はストレスに曝されると成長を止めますが、これは植物が能動的に成長を抑制して、節約したエネルギーをストレス対処に使っている表れです。このような「エネルギーの再配分」は植物が変動する環境下で生きていく上で非常に大切ですが、そのメカニズムは不明のままでした。
私達は、DNA損傷や高温などの複数のストレスに応答して細胞分裂を停止させるシグナル伝達経路を発見しました。転写因子ANAC044, ANAC085を介して細胞周期をG2期で停止させる経路です。これはまさにストレス下でのエネルギー再配分に働く、植物にとって重要な環境応答機構の一つと考えられますが、人間が栄養分を操作できる農地では作物の生産性を下げるマイナス要因となります。逆に言えば、このシグナル伝達経路を抑制することで収量増加につながると期待されます。そこで、私達はこの制御システムの全容解明に取り組むことで、環境ストレスに曝されても成長を止めない作物の育種に貢献したいと考えています。

図2. DNA損傷や高温ストレスは転写因子ANAC044, ANAC085の発現を誘導し、別の転写因子であるMYB3R3, MYB3R5の蓄積量を上げることを見出している。これにより細胞周期がG2期で停止し、成長が止まる。このシグナル伝達経路を抑制することで、ストレス下でも成長を止めない作物の創出が期待できる。
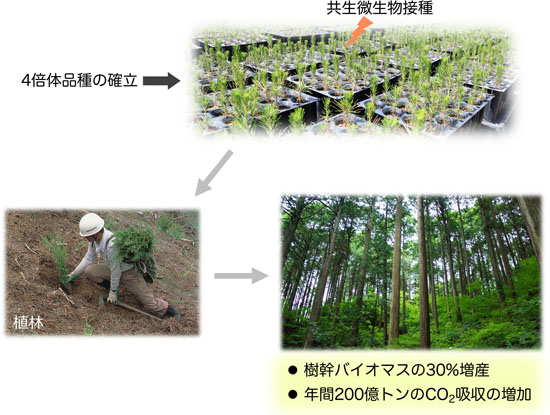
3. 樹幹バイオマスの増産技術の開発
植DNA倍加は一つの細胞の中でゲノムDNA量が倍々に増えていく現象です。動物では稀にしか見られませんが、植物では約70%の種で起こる現象で、細胞・器官サイズを増大させる効果をもっています。そのため、数多くの農作物の育種に利用されてきました。しかし、樹木ではDNA倍加が全く起きません。その理由は未だ不明ですが、樹木でもDNA倍加により器官サイズを大きくすることができれば、木質バイオマスの増産や二酸化炭素吸収の増大につながると考えられます。
私達は以前、樹木でDNA倍加を誘発する技術開発に取り組みました。その結果、DNA量が倍になった4倍体ポプラを作出することに成功しましたが、このポプラは幹が太くなる一方で縦方向の伸長が遅くなり、結果的に樹幹バイオマスは元の2倍体と変わらない、という結果が得られました。もしも縦方向の伸長が回復すれば、樹幹バイオマス生産量は30%増えると見込んでいます。
そこで私達は現在、樹木の伸長を促進する共生微生物の探索を行っています。4倍体化した樹木種に共生微生物を接種し、それを植林して育てれば、DNA倍加と共生微生物による縦横両方向の成長促進効果を持続させることができるので、樹幹バイオマスの飛躍的増産につながると考えています。また、私達は非DNA倍加植物の4倍体化を効率的に実現するために、DNA倍加の誘導機構についてもエピジェネティックな視点から解析を進めています。これらの研究を通して、地球レベルのカーボンリサイクルの実現に貢献したいと考えています。