
2024.04.01
高温環境下における分裂酵母の生育抑制メカニズム
微生物インタラクション研究室・助教・両角 佑一
- 要旨
- 高温環境では、細胞を構成するタンパク質などの熱損傷によって細胞増殖は停止すると考えられています。最近、私達はTORC1キナーゼ複合体の阻害剤であるラパマイシンによって、分裂酵母が通常では生育できない高温でも生育できるようになることを発見しました。このことは、細胞増殖を促進因子として知られるTORC1が、高温では増殖を抑制していることを示唆しています。さらに、この発見を足がかりとして、分裂酵母にはこれまで考えられていたよりも高い温度で増殖できる能力が備わっているものの、その能力はTORC1をはじめとする複数の因子によってあえて抑えられていることを明らかにしました。本稿では、高温環境における分裂酵母の細胞増殖抑制メカニズムについて紹介します。
- 主要関連論文
- Morozumi Y, Mahayot F, Nakase Y, Soong JX, Yamawaki S, Sofyantoro F, Imabata Y, Oda AH, Tamura M, Kofuji S, Akikusa Y, Shibatani A, Ohta K, Shiozaki K. Rapamycin-sensitive mechanisms confine the growth of fission yeast below the temperatures detrimental to cell physiology. iScience, 2024, 27, 108777.
doi:10.1016/j.isci.2023.108777
1.はじめに
地球上で生物が生息する環境温度は、下は氷点下から上は100℃以上と驚くほど多様ですが、個々の生物種が生育できる温度の範囲は比較的狭く、その範囲からわずかに温度が上昇するだけで生存を脅かす熱ストレスになりえます。熱ストレスによって生じるタンパク質の変性や凝集を防ぐために、細胞は進化的に保存された熱ショック反応を活性化し、分子シャペロンとして機能する熱ショックタンパク質 (Heat Shock proteins: HSPs)の発現を誘導します1。これまで、様々な生物種をモデルとしてHSPsに関する研究が盛んに行われ、熱ストレスに対する細胞の生存戦略が明らかになってきた一方で、持続した高温環境での細胞の成長・増殖に関する研究はあまり進んでいないのが実情です。
分裂酵母の至適生育温度は30℃程度であり、38℃を超えると生育できずに死に至ります。しかし、放線菌が産出するマイロライド化合物であるラパマイシンを培地に添加することで、分裂酵母が39℃の高温でも増殖できるようになることを私達は発見しました(図1)。現在、私たちはこの発見を足がかりとして高温環境下での細胞増殖制御メカニズムに興味を持って研究を進めています。
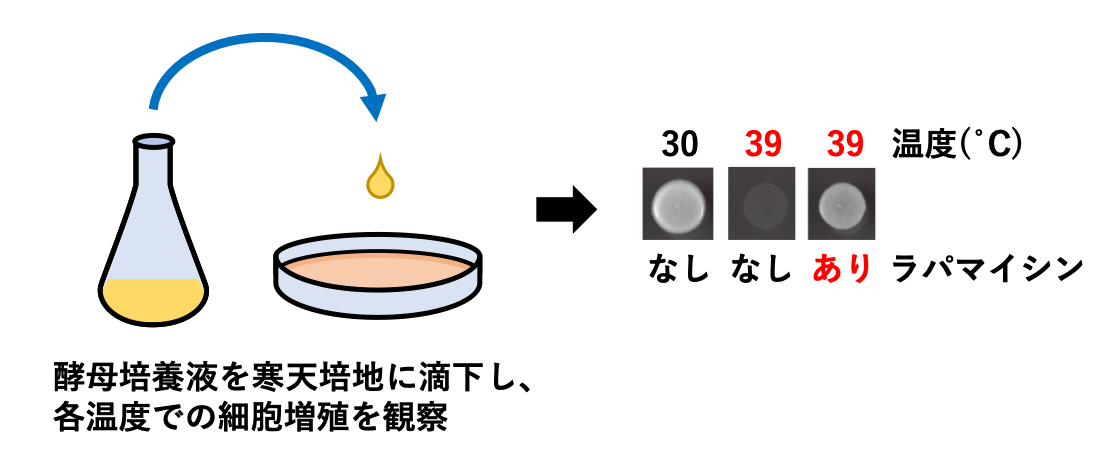
図1.ラパマイシンによって分裂酵母は高温で生育できるようになる
2.TORC1シグナル伝達経路を介した細胞増殖の抑制
ラパマイシンの細胞内標的因子は、TOR (Target of Rapamycin)が形成するキナーゼ複合体TOR complex 1 (TORC1)であり、TORC1は栄養源に応答し細胞の成長・増殖の促進するシグナル伝達経路で中心的な役割を果たすことが知られています2,3。ラパマイシンは細胞内でFKBP12タンパク質と結合し、その“FKPB12-ラパマイシン複合体”がTORC1に結合することで活性を阻害します。そこで、FKBP12タンパク質をコードするfkh1遺伝子を欠失し、ラパマイシンによるTORC1阻害が消失した分裂酵母株の高温での生育を観察した結果、予想通りラパマイシン存在下でも生育できませんでした。また、TORC1の制御サブユニットであるMip1に変異を導入することでTORC1の活性が低下した変異株4は、ラパマイシンがなくても高温で生育できることがわかりました。これらの結果から、増殖促進因子として知られているTORC1が高温環境では増殖を抑えていることが明らかになりました(図2)。
TORC1は、細胞内で様々な基質をリン酸化することで細胞成長・増殖を制御します。分裂酵母では、AGCキナーゼファミリーに属するPsk1、Sck1、Sck2やRNAポリメラーゼIIIの抑制因子であるMaf1、さらにはオートファジー制御因子であるAtg13などがTORC1の基質として報告されています5。そこで、これら基質の遺伝子破壊株の高温生育を解析したところ、sck1遺伝子破壊株のみが39℃でラパマイシン非存在下でも生育を示しました。これらの結果から、TORC1は下流のSck1を介して高温生育を抑制することが示唆されました。
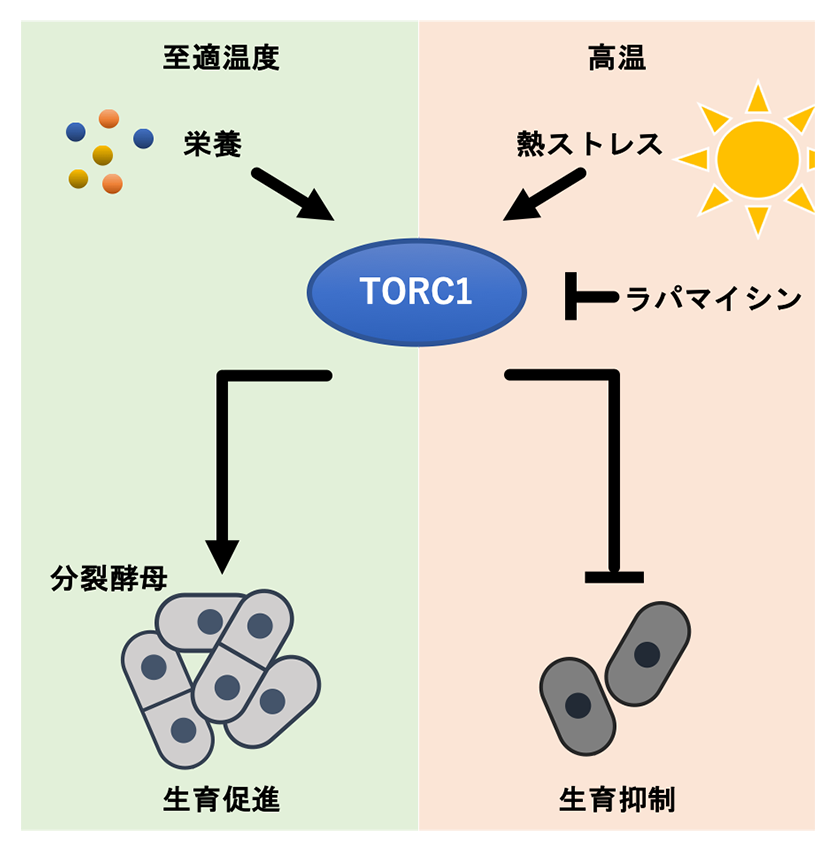
図2.TORC1による生育制御
至適温度ではTORC1は栄養に応答して生育を促進する一方で、高温環境下では生育を抑制する。
3.高温生育を抑制する因子の網羅的探索と関連の解析
Sck1はキナーゼであり、下流の基質をリン酸化することで高温生育を抑制すると考えられますが、その基質は未だに不明です。また、Sck1以外にも高温生育の抑制因子が存在することが予想されます。そこで、分裂酵母の非必須遺伝子が1つずつ欠失した遺伝子破壊株ライブラリーを用いて、ラパマイシンがなくても高温生育を示す変異株をスクリーニングした結果、Sck1に加えてMks1、Dri1およびRhs1が高温増殖を抑える機能を持つ因子として同定されました。また、野生型株を39℃に数日間おくことで高温での増殖能を獲得した突然変異株が得られたため、変異株に含まれる遺伝子変異を特定し、高温生育抑制因子をさらに探索した結果、14-3-3タンパク質であるRad24が同定されました。これらの因子の関連についてさらに解析を進めたところ、Mks1はRad24と、Dri1はRhs1と複合体を形成して機能することが明らかになりました。さらに、ラパマイシンによってMks1のリン酸化が抑えられることや、Mks1とTORC1の相互作用が検出されたことから、Mks1はTORC1の基質であることがわかり、同じくTORC1の下流で機能するSck1とは別経路で高温での生育を抑制していることも見出しました。これらのことから、分裂酵母にはこれまで考えられていたよりも高い温度で増殖できる能力が備わっているものの、TORC1をはじめとする複数の因子によってあえて抑制されていることが示唆されました(図3)。
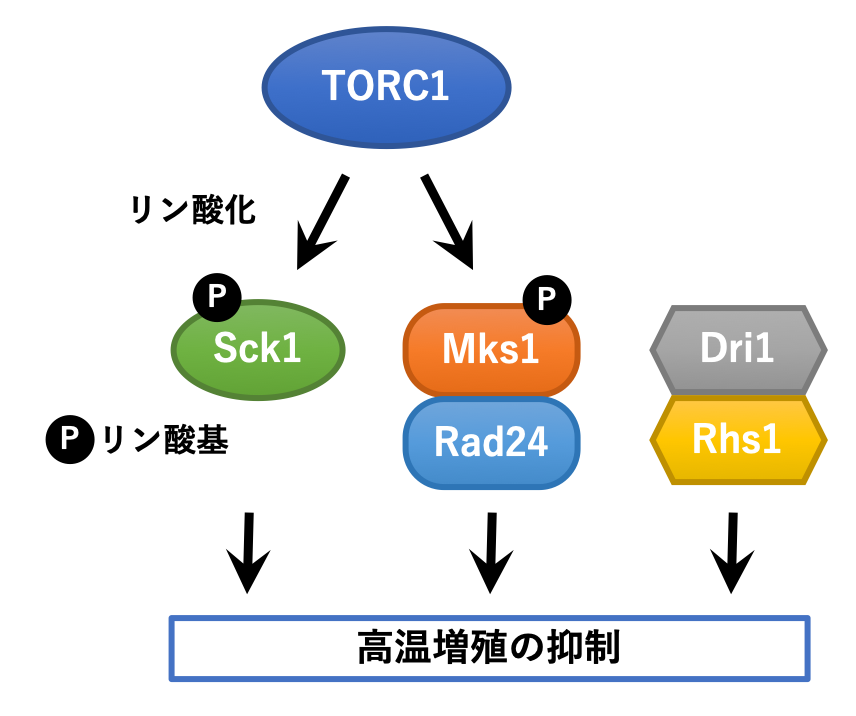
図3.分裂酵母の高温生育の抑制経路
Mks1はRad24と、Dri1はRhs1と相互作用して機能する。また、Sck1とMks1はTORC1の下流で、互いに独立して生育を抑制する。
4.おわりに
高温環境では細胞を構成するタンパク質などの熱損傷によって増殖が停止すると考えられていますが、私達の発見は、分裂酵母の増殖温度の上限が細胞の熱損傷の程度ではなく、細胞内の制御メカニズムによって決定されている可能性を示しています。地球温暖化の進行に伴い、ヒトを含むさまざまな生物種が高温ストレスに晒される危険性が年々増しているため、高温環境に生物がどのように対応しているのかを理解することはとても重要です。興味深いことに、本研究で発見した高温増殖を抑制する因子の多くはヒトにも存在しています。今後、分裂酵母で見つかった高温生育の抑制メカニズムをヒトでも明らかにできれば、熱ストレス緩和方法の開発などにつながることも期待できます。
参考文献

著者
両角 佑一 Researchmap
略歴
2011年早稲田大学大学院先進理工学研究科博士課程修了後、早稲田大学先進理工学部助手、同助教、フランス Institut Albert Bonniot 博士研究員などを経て、2017年より現職
両角 佑一 NAIST Edge BIO, 0023. (2024)