
2022.07.01
DNA損傷に対する植物の応答とその制御機構
植物成長制御研究室・助教・髙橋直紀
- 要旨
- 自由に移動することができる動物とは異なり、陸上植物は一度根付いた場所から動くことが出来ません。そのため、植物は周囲の環境変動に適応しながら成長をコントロールする必要があります。遺伝情報であるDNAは、通常の複製過程でエラーを生じ、それが原因となり損傷を受けます。また、環境ストレスもDNAに損傷を与えることがあります。近年の研究により、植物はDNA損傷応答に働くユニークな制御因子を持つことが明らかになってきました。本稿では、植物のDNA損傷応答について概説するとともに、最近明らかになった植物独自の分子メカニズムについて紹介します。
1.はじめに
二重らせん構造をもつDNAは、アデニン(A)、チミン(T)、グアニン(G)、シトシン(C)の4つの塩基から構成されています。そして、すべての生物の体は、これら塩基配列の情報に基づいて作られます。そのため、もしDNAが損傷を受けて塩基配列に変異が入ってしまうと、動物ではがんをはじめとする広範な疾患の原因となるなど、全ての生物において生存に影響を与える危険性があります。
一方で、DNAは常に損傷の危機に晒されていることが知られています。例えば、通常のDNA複製の過程で生じる複製エラーや、細胞内で発生する活性酸素によりDNAは損傷を受けます。また、紫外線や放射線によってもDNA損傷を受けることや、植物では土壌中に含まれる過剰なアルミニウムやホウ素、病原菌感染などによっても損傷を受けることが示されています。動物には、p53などの制御因子を介したDNA損傷応答機構が存在し、DNA修復や細胞周期停止、アポトーシスなどを制御していることが知られています。このような応答機構が働くことで、軽微なDNA損傷は修復され、致命的なDNA損傷に対しては変異が入ったDNAが娘細胞へ伝わるのを防ぐため、細胞を積極的に死滅させます。植物も同様にDNAに損傷が与えられると、DNA修復や細胞周期停止などが起こることが知られていますが、幹細胞特異的な細胞死やDNA倍加(細胞分裂をせずにDNA複製を繰り返す細胞周期で、核DNA量を増加させることで細胞を肥大化させる)の誘導など、他の生物で見られないような植物独自の応答機構の存在も明らかにされつつあります。本稿では、進化の過程で獲得してきた植物独自のDNA損傷応答経路について紹介します。
2.植物のDNA損傷応答
DNAが損傷を受けると、真核生物ではATMとATRという2つのセンサーキナーゼが損傷部位に結合します。動物や酵母の場合、損傷部位に結合したATMやATRはエフェクターキナーゼであるCHK1、CHK2をリン酸化し、活性化します。さらに、動物細胞の場合、p53、CDC25、53BP1などの因子がATM、ATR、CHK1、CHK2により制御されることで、DNA修復や細胞周期停止、アポトーシスなどを制御しています。同様に、植物もATM、ATRが損傷DNAの認識に関わっていることが知られています。しかし、植物はCHK1やCHK2、p53、53BP1のような動物のDNA損傷応答に重要な因子のオルソログをもっていません。一方で、植物特異的な転写因子SOG1がATM、ATRによりリン酸化されることで活性化し、DNA修復、CDK活性の低下、病原菌感染応答、植物ホルモン応答などに関わる下流因子の発現を誘導していることが明らかにされています(図1)1)2)。そして、SOG1 を欠損した変異体では、細胞周期停止などのDNA損傷応答がほとんど起きなくなることから3)、SOG1転写因子が植物のDNA損傷応答において重要な役割を果たしていると考えられています。
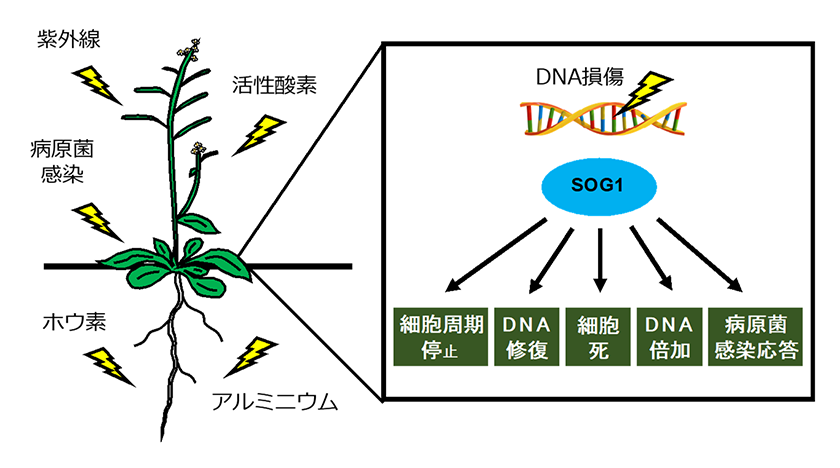
図1.植物のDNA損傷応答
植物はDNA損傷を受けると、転写因子SOG1が活性化することで細胞周期の停止、DNA修復、幹細胞の細胞死、DNA倍加の誘導、病原菌感染応答などのDNA損傷応答を引き起こす。
3.細胞周期の停止
生物はDNAに損傷を受けると、細胞周期チェックポイント機構が働くことで細胞周期を一時的に停止(もしくは、遅延)させ、損傷箇所の修復を行います。そのことから、DNA損傷に応答した細胞周期の停止は、ゲノムの恒常性維持に非常に重要な機構になります。そして、DNA損傷に晒されると細胞周期進行を制御するサイクリン依存性キナーゼ(CDK)の活性が低下することで、細胞周期の停止が起きることが知られています。植物においても、DNA損傷が起きるとCDKインヒビターであるSMR の遺伝子の発現がSOG1依存的に上昇することで、CDK活性の低下が起きていることが示唆されています4)。
これまでに植物成長制御研究室では、M期サイクリンなどのG2/M期特異的遺伝子群の発現抑制に関わる3R型MYB転写因子(Rep-MYB)の蓄積がDNA損傷に応答したG2期での細胞周期停止に関与していることを明らかにしていました5)。Rep-MYBタンパク質はストレスを受けていない時にはCDKによりリン酸化されることでプロテアソームにより分解されていますが、DNA損傷によりCDK活性が低下するとRep-MYBタンパク質のリン酸化レベルが低下することで分解されずに蓄積します。そして、Rep-MYBタンパク質が蓄積することでG2/M期特異的遺伝子群の発現が抑制されて、G2期停止が引き起こされます。また、最近の研究で、CDK活性の低下に加えて、SOG1の標的因子であるNAC型転写因子のANAC044とANAC085もRep-MYBタンパク質の安定化に関与していることが明らかになっています6)。このことから、DNA損傷を受けた際に、植物は複数の経路を介して細胞周期をG2期で停止させていると考えられます(図2)。
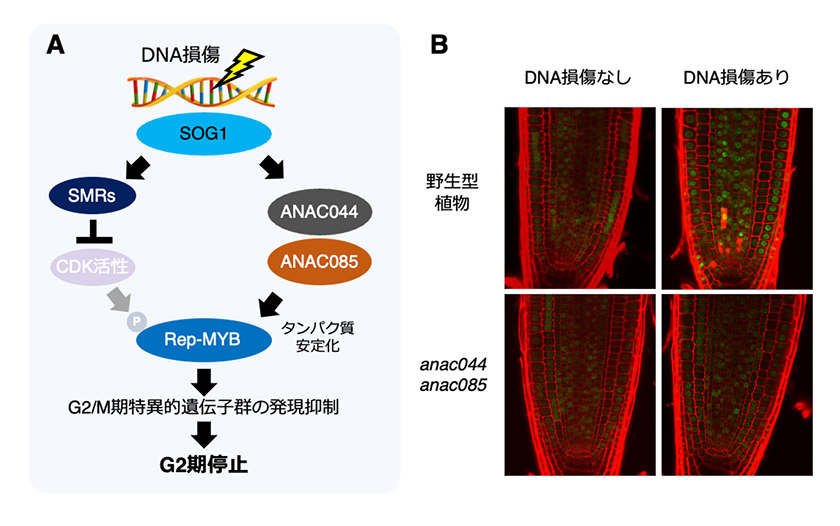
図2.DNA損傷に応答したG2期での細胞周期停止機構
(A) DNA損傷が起こると、SOG1はCDKインヒビターであるSMRs の発現を誘導し、CDK活性を低下させる。また、ANAC044/085 の発現も上げることで、Rep-MYBタンパク質を安定化させる。 Rep-MYBタンパク質が蓄積するとG2/M期特異的遺伝子群の発現が抑制され、G2期で細胞周期が停止する。
(B) DNA損傷時のRep-MYB(MYB3R3)タンパク質の蓄積。 DNA二本鎖切断を引き起こすゼオシンでシロイヌナズナの根を処理した後、pMYB3R3:MYB3R3-GFPM のGFPシグナルを観察した。野生型植物では、DNA損傷によるMYB3R3-GFPの蓄積が起こるが、anac044 anac085変異体では起こらない。
興味深いことに、Rep-MYBタンパク質は、高温ストレスに晒された時にも高蓄積することで細胞周期停止に関わりますが、高温ストレス時のRep-MYBタンパク質の蓄積にもANAC044/085が関与していることが明らかになっています6)。このことから、ANAC044/085は様々な環境ストレスに応答して細胞周期をG2期で停止させるのに機能している可能性が考えられます。一方で、ANAC044/085がどのようにRep-MYBタンパク質の安定化に寄与しているのかについては未だ明らかにされていません。今後は、Rep-MYBタンパク質の安定化機構を明らかにすることで、ANAC044/085を介した細胞周期停止機構の理解がさらに進むことが期待されます。
4.幹細胞の細胞死
DNA損傷が起きると、軽微な損傷の場合は速やかに修復しますが、修復不能な重篤な損傷の場合には細胞死が誘導されます。植物では、DNA損傷を受けると、すべての細胞で細胞死が誘導されるのではなく、幹細胞でのみ細胞死が引き起こされることが知られています7)。幹細胞は様々な組織、器官を構成する細胞を生み出すことから、植物ではDNA損傷により幹細胞の染色体DNAが傷つくと、それが娘細胞に伝わらないようにするために、積極的に幹細胞で細胞死を引き起こしているのかもしれません。ATMやATR、SOG1を欠損したシロイヌナズナ変異株では、DNA損傷後の幹細胞の細胞死が起きないことから、DNA損傷応答経路を介して積極的に幹細胞の細胞死が誘導されていることが考えられます。最近の研究で、私達はシロイヌナズナをDNA損傷誘導剤で処理すると、根端で植物ホルモンのオーキシンが低下することを発見しました。また、DNA損傷誘導剤と同時にオーキシンを処理すると、幹細胞の細胞死が抑制されることも明らかにしています8)。このことから、幹細胞の細胞死にオーキシンの低下が深く関与していると予想されます。今後は、オーキシンの低下がどのように幹細胞特異的な細胞死を誘導しているか明らかにすることにより、植物独自のホルモンによるゲノム恒常性の維持機構に迫ることができると考えています。
5.DNA倍加の誘導
当研究室では、シロイヌナズナがDNA損傷を受けると、通常の細胞周期からDNA倍加周期への移行が早まることを明らかにしています9)。つまり、DNA損傷に晒されると、細胞分裂は停止しながらもDNA倍加を誘導することで細胞伸長を促進し、器官成長をある程度保証すると考えられます。DNA倍加への移行にはE3ユビキチンリガーゼである後期促進複合体APC/Cの活性化因子であるCCS52A1 の発現誘導が関わっていることが知られています。そして、シロイヌナズナの根では、植物ホルモンのサイトカイニンのシグナル伝達がCCS52A1 の転写誘導に直接関わっていることが明らかにされていました10)。最近の私達の研究により、DNA損傷を受けるとサイトカイニンの生合成が活性化することで、根端でサイトカイニンが高蓄積することを発見しました。そして、サイトカイニンシグナルの活性化によりCCS52A1 遺伝子の発現が誘導され、DNA倍加への移行が促進されることを見出しました。さらに、サイトカイニンシグナルが活性化すると、オーキシンシグナルの抑制に関わるSHY2 の発現が誘導されることで、根の基部側から根端側へのオーキシン輸送に関わるPIN の発現を低下させることを明らかにしました。そして、オーキシン輸送が阻害されることで根端でのオーキシンレベルが低下し、それが細胞周期の停止、幹細胞特異的な細胞死の誘導に関わることを発見しました(図3)8)。このことから、シロイヌナズナではサイトカイニンとオーキシンの量およびシグナルを変化させることにより、DNA倍加の誘導に加えて、細胞周期の停止、幹細胞の細胞死などの、根での様々なDNA損傷応答が統御されていることが考えられます。一方で、他の植物ホルモンもDNA損傷応答に関わっていることも示唆されています。今後は、DNA損傷時の植物ホルモン間のクロストークを解明することで、根でのDNA損傷応答機構がより詳細に明らかになることが期待されます。
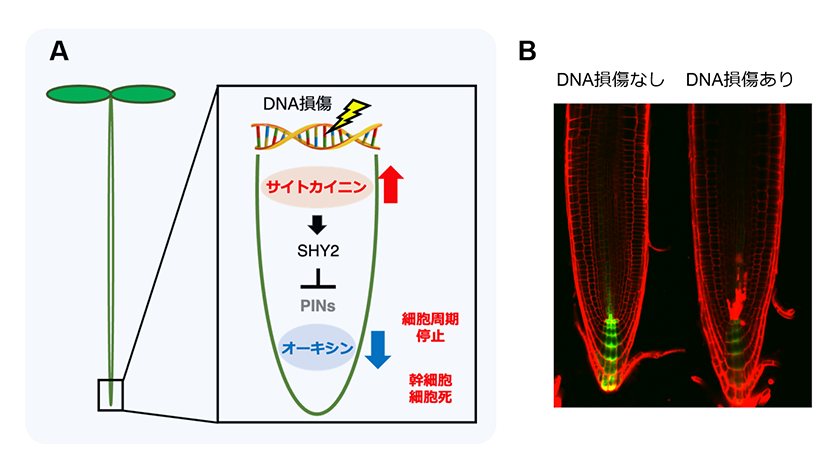
図3.サイトカイニン、オーキシンによるDNA損傷応答制御
(A) DNA損傷を受けると、シロイヌナズナの根端ではサイトカイニンが蓄積する。サイトカイニンサイトカイニンシグナルは、SHY2 の発現を誘導し、PIN の発現を抑制することによりオーキシンシグナルを低下させる。オーキシンシグナルの低下は、細胞周期の停止、幹細胞の細胞死を誘導する。
(B) DNA損傷後の根端でのオーキシンシグナル変化。DNA二本鎖切断を引き起こすゼオシンでシロイヌナズナの根を処理した後、オーキシン応答マーカー(DR5rev:GFP )を観察した。DNA損傷後にオーキシンシグナルが低下する。
6.おわりに
植物のDNA損傷応答において重要な役割を果たしているSOG1転写因子の発見により、植物のDNA損傷応答の実像が徐々に解明されつつあります。そして、上記の通り、植物特異的な転写因子ANAC044/085や、植物ホルモンのサイトカイニン、オーキシンが、DNA損傷に応答した細胞周期の停止や幹細胞の細胞死、DNA倍加の誘導に関わることが明らかになってきました。また、病原菌の感染応答にもDNA損傷応答経路が重要な役割を果たしていることも示唆されていることから、植物は他の真核生物とは異なる独自のDNA損傷応答経路を進化の過程で獲得してきたと考えられます。今後、植物のDNA損傷応答の理解がさらに進めば、動物や酵母とのDNA損傷応答との類似性や相違点などが明らかにされるとともに、植物独自のユニークな制御メカニズムがさらに明らかになることが期待されます。
参考文献
- Ogita, N. et al., Plant J.(2018).
- Bourbousse, C. et al., PNAS.(2018).
- Yoshiyama, K. et al, PNAS.(2009).
- Yi, D. et al., Plant Cell.(2014).
- Chen, P. et al., Nat Commun.(2017).
- Takahashi, N. et al., eLife.(2019).
- Fulcher, N. & Sablowski, R. PNAS.(2009).
- Takahashi, N. et al., Sci Adv.(2021).
- Adachi, S. et al., PNAS(2011).
- Takahashi, N. et al., Curr Biol.(2013)

著者
高橋 直紀 Researchmap
略歴
2006年 東京理科大学大学院博士課程修了、同年 ベルギー・ゲント大学 博士研究員
2009年 理化学研究所 特別研究員、
2011年 奈良先端科学技術大学院大学 博士研究員を経て、2013年より現職。
- 研究内容:植物のストレス応答機構
- 抱負:常に新しい発見をする
- 関心ごと:世界情勢
高橋 直紀 NAIST Edge BIO, 0002. (2022)