Research 研究成果の紹介
酵母が環境に合わせて発酵力を変える仕組みを解明 自在に能力を高める「発酵デザイン技術」の確立へ ~清酒酵母の高発酵力の原因も明らかに~
酵母が環境に合わせて発酵力を変える仕組みを解明
自在に能力を高める「発酵デザイン技術」の確立へ
~ 清酒酵母の高発酵力の原因も明らかに ~
【概要】
奈良先端科学技術大学院大学(学長:横矢直和)先端科学技術研究科バイオサイエンス領域ストレス微生物科学研究室の渡辺大輔助教、高木博史教授の研究グループらは、酒づくりの過程で酵母が発酵する際に、糖など環境中の栄養状態を感知し、そのシグナル(情報)を伝達して発酵力を変えるという重要な仕組みを初めて解明しました。さらに、日本特有の清酒酵母が本来持っている高発酵力を生み出すメカニズムとして、発酵の初期の段階で、発酵のブレーキ役の酵素の働きが抑制され、エンジン役となる酵素の複合体が活性化されていることも明らかにしました。これで、酒やパンなどの発酵技術で酵母の発酵力を自在に改変する「発酵デザイン技術」に結びつくことが期待できます。
酵母は、アルコール発酵によって糖類からエタノールと二酸化炭素を生成し、酒類、パン、バイオエタノールなどの製造に不可欠な微生物です。ところが、酵母の発酵力を人為的に改変することは容易ではなく、発酵産業において解決が望まれていました。一方、発酵産業において用いられる実用酵母菌株には、高い発酵力を有するものが多数存在しますが、その原因についてもほとんど明らかにされていませんでした。そこで、本研究では、我が国独自の微生物資源であり、実用酵母菌株の中でも高い発酵力を有する清酒酵母に着目することで、酵母が発酵力を調節するメカニズムの理解を目指しました。
酵母は、環境中の栄養源に応答して発酵力を変化させることが知られています。このことから、真核生物(核を持つ生物)に広く保存された、栄養状態を感知し伝達する調節システムの主役として知られるプロテインキナーゼ(タンパク質リン酸化酵素)の複合体(TORC1)に着目しました。その結果、TORC1を活性化させると発酵の立ち上がりの勢いが強くなる現象を見出しました。さらに、TORC1に続いて働く発酵のブレーキ役のGreatwall(グレイトウォール)というプロテインキナーゼと、エンジン役のプロテインフォスファターゼ(タンパク質加水分解酵素)の複合体(PP2AB55δ)も発酵力の調節に必須であることを明らかにしました。清酒酵母でこのシグナル伝達経路を解析したところ、発酵初期においてTORC1活性が高く、Greatwallの機能が欠損していたことから、PP2AB55δが強化されている可能性が示唆されました。実際に、清酒酵母においてPP2AB55δを欠損させると発酵力が顕著に低下したことから、PP2AB55δが清酒酵母の高い発酵力を生み出す原因であることが証明されました。
本研究で得られた知見を応用することで、実用酵母菌株の発酵力を自在に改変することが可能となり、発酵産業にとって有用な「発酵デザイン技術」の確立が期待されます。また、TORC1、Greatwall、PP2AB55δは、いずれも真核生物に高度に保存されていることから、より高等な生物における炭素代謝調節メカニズムの解明にも貢献できると考えられます。
この研究成果は、米国微生物学会の学会誌であるApplied and Environmental Microbiology誌オンラインサイトに平成30年10月19日付で掲載されました。
【渡辺大輔助教からのコメント】
お酒づくりに使われる「清酒酵母」はアルコール発酵力が高いことで知られていますが、その原因は長年にわたり謎とされてきました。今回の論文では、真核生物に保存されたTORC1、Greatwall、PP2AB55δというシグナル伝達因子が、酵母によるアルコール発酵の調節に重要な役割を果たしていること、そして、清酒酵母はこれらの因子に違いが生じたことで高いアルコール発酵力を獲得したことを明らかにすることができました。今後、この清酒酵母での知見を生かして、他の実用酵母菌株のアルコール発酵力を自在に改変することも可能になるのではないかと期待されます。ちょうどお酒がおいしくなってくるこの時期に、清酒酵母という、私たちの生活に身近な生き物の謎を一つ解明することができて、大変うれしく思います。これからも、「発酵」という奥の深い生命現象をとことん研究して、科学的に面白い発見につなげていくことができればと思います。
本研究は主として、バイオサイエンス領域ストレス微生物科学研究室において実施されました。高木博史教授には、本研究に集中的に取り組むための環境を与えていただいたことに加え、酵母・発酵研究の重要性と楽しさを熱心にご指導いただいております。同研究室の修了生である梶原拓真君、高木健一君はこの論文の完成につながる新しい発見を成し遂げてくれましたし、研究技術員の杉本幸子さんには研究期間を通してたえず献身的なサポートをいただきました。この場を借りまして、心より感謝申し上げます。最後に、本研究は、学内外の多くの酵母研究者の皆様方との共同研究により行われたものであり、ご指導とご支援に深謝申し上げます。

【解説】
〈研究背景〉
酵母によるアルコール発酵については、微生物学・生化学の黎明期から多くの研究がなされ、必要な酵素・遺伝子がほぼ完全に明らかにされています。それにも関わらず、現代のバイオテクノロジーをもってしても、「ある酵母菌株の発酵力を高めるにはどうすれば良いか?」という素朴な疑問に答えることが容易ではありません。その理由は、酵母が有する発酵調節メカニズムが未解明であったからです。したがって、酒類、パン、バイオエタノールなどの製造に用いられる実用酵母菌株を育種するためには、多くの菌株を用いて発酵試験を行い、実際に高い発酵力を示す菌株を選抜するという、時間と労力を必要とする手法が用いられています。そこで、発酵調節メカニズムを解明し、その知見を応用することによって効率良く発酵力を改変する育種技術の確立が望まれています。発酵調節メカニズムの解明に大きなヒントを与えてくれるかもしれないのが、高い発酵力を有する実用酵母菌株です。中でも、世界で最もアルコール度数の高い醸造酒である清酒の製造に用いられる清酒酵母は、優れた発酵力を有することで知られていますが、その原因についても未解明のままでした。
酵母は、環境中の栄養源に応答して発酵力を変化させることが知られており、栄養源が豊富な環境では旺盛に発酵を行うのに対し、何らかの栄養源が枯渇すると発酵は阻害されます。したがって、環境中の栄養状態をインプットとして、発酵力というアウトプットを生み出す栄養シグナリング経路が、発酵調節メカニズムに関与しているのではないかと推測されます。酵母からヒトに至るあらゆる真核生物において、TORC1と呼ばれるプロテインキナーゼ複合体が栄養シグナリングのマスターレギュレーターとして機能することが知られており、細胞の成長や増殖、遺伝子発現、オートファジー、寿命など多くの生命現象に関与しています。しかしながら、現在までにアルコール発酵への影響は解析されていませんでした。そこで本研究では、発酵調節におけるTORC1を介した栄養シグナリング経路の役割を明らかにし、清酒酵母の高発酵力を生み出す原因を解明することを目的としました。
〈研究結果〉
本研究では、グルコースを炭素源とする培地において発酵試験を行い、発酵に伴い生じる二酸化炭素の量を測定することで、発酵力の解析を行いました。まず、出芽酵母Saccharomyces cerevisiae(サッカロミセス・セレビシエ)に属する実験室酵母をモデルとして用い、TORC1の活性を改変したところ、発酵初期における発酵速度に変化が生じることがわかりました(図1A, B)。特に、TORC1を活性化させると発酵の立ち上がりの勢いが強くなる現象が見出されました。このことから、TORC1活性と発酵力との密接な関係が明らかになりました。さらに、TORC1の下流で働く標的因子のうち、GreatwallをコードするRIM15遺伝子を破壊すると発酵速度が上昇し(図1C)、PP2AB55δの触媒サブユニットをコードするCDC55遺伝子を破壊すると発酵速度が低下したことから(図1D)、Greatwallが発酵の「ブレーキ」、PP2AB55δが発酵の「エンジン」として働くことが示されました。このようなTORC1-Greatwall-PP2AB55δ経路と発酵力との関係は、Saccharomyces cerevisiaeとは進化的にかけ離れた種である分裂酵母Schizosaccharomyces pombe(シゾサッカロミセス・ポンべ)においても成り立っていたことから、真核生物に共通の代謝調節経路が形成されている可能性が高いと考えられます。次に、細胞内の代謝物を網羅的に調べるメタボローム解析※5の結果、実験室酵母のCDC55遺伝子破壊株では、アルコール発酵の中間代謝産物であるフルクトース-6-リン酸(F6P)が蓄積し、フルクトース-1,6-ビスリン酸(F1,6BP)の含有量が低下していました(図2)。このことから、PP2AB55δが、F6PからF1,6BPへの変換を触媒するフォスフォフルクトキナーゼ(PFK)の活性を正に制御することで発酵力を高めるというメカニズムも明らかになりました。清酒酵母でこのTORC1-Greatwall-PP2AB55δ経路を解析した結果、発酵初期におけるTORC1活性が高く(図3)、Greatwallの遺伝子であるRIM15遺伝子上に機能欠失変異を有していました。清酒酵母においてCDC55遺伝子を破壊すると発酵速度が顕著に低下したことから(図1E)、高いTORC1活性とGreatwallの欠損によってPP2AB55δの機能が強化されたことが、清酒酵母の高発酵力を生み出す原因であることが証明されました。
<研究意義>
本研究により、真核生物に保存されたTORC1-Greatwall-PP2AB55δ経路を介する栄養シグナリングが酵母の発酵力調節のための鍵を握っており(図4)、中でも、発酵の「エンジン」であるPP2AB55δが清酒酵母の高発酵力に必須であることが解明されました。従来、「発酵力の低下を引き起こすのは酵母の死滅であり、酵母の生存率を高めれば発酵力を改善できる」と考えられてきました。しかしながら、実際は、生きている酵母はTORC1によって栄養状態をモニターしながら発酵力をダイナミックに変化させる能力を持つことが示されました。そして、発酵を阻害するメカニズムを欠損させることによって清酒酵母のような高い発酵力を有する菌株を効率良く獲得することができることもわかりました。今回の研究で得られた知見は、酒類、パン、バイオエタノールなどの製造に用いられる実用酵母菌株の発酵力を高めることにも応用可能であり、発酵産業にとって有用な「発酵デザイン技術」の確立につながることが期待されます。さらに、TORC1、Greatwall、PP2AB55δは、いずれも真核生物に高度に保存されていることから、より高等な生物における炭素代謝調節メカニズムの解明にも貢献できると考えられます。
【本研究について】
本研究は、奈良先端科学技術大学院大学、酒類総合研究所、東京大学、甲南大学、秋田県立大学、京都工芸繊維大学、岩手大学との共同研究により実施されました。
【掲載論文】
タイトル:Nutrient signaling via the TORC1-Greatwall-PP2AB55δ pathway responsible for the high initial rates of alcoholic fermentation in sake yeast Strains of Saccharomyces cerevisiae
(和訳:TORC1-Greatwall-PP2AB55δ経路を介した栄養シグナリングが清酒酵母の高いアルコール発酵力を生み出している)
DOI : https://doi.org/10.1128/AEM.02083-18
著者:Daisuke Watanabe1,2), Takuma Kajihara1), Yukiko Sugimoto1), Kenichi Takagi1), Megumi Mizuno2), Yan Zhou2), Jiawen Chen3), Kojiro Takeda4,5), Hisashi Tatebe1), Kazuhiro Shiozaki1), Nobushige Nakazawa6), Shingo Izawa7), Takeshi Akao2), Hitoshi Shimoi2,8), Tatsuya Maeda3), Hiroshi Takagi1)
所属:1)奈良先端科学技術大学院大学先端科学技術研究科、2)酒類総合研究所、3)東京大学分子細胞生物学研究所, 4)甲南大学理工学部、5)甲南大学統合ニューロバイオロジー研究所、6)秋田県立大学生物資源科学部、7)京都工芸繊維大学大学院工芸科学研究科、8)岩手大学農学部
掲載誌:Applied and Environmental Microbiology
naister:http://hdl.handle.net/10061/12605(NAIST Academic Repository:naistar)
【用語解説】
※1:酵母
単細胞性の真菌類の総称であり多くの種を含むが、中でも最も有名なのが、発酵産業に広く用いられている出芽酵母Saccharomyces cerevisiaeである。この種には、基礎研究のモデル株として用いられる実験室酵母や、清酒醸造に用いられる清酒酵母、パンの製造に用いられるパン酵母などが含まれ、それぞれの用途に応じた適性を有している。例えば、実験室酵母は、遺伝解析やゲノム解析による知見が蓄積され遺伝子操作技術も確立しているが、発酵力は弱く発酵食品などの生産には不適である。一方、清酒酵母やパン酵母などの実用酵母菌株は、発酵力が高く、各発酵食品を特徴づける香味成分の生成能に優れている。Saccharomyces cerevisiaeが出芽によって増殖するのに対し、分裂によって増殖するSchizosaccharomyces pombeなどの種も存在し、基礎研究のモデル株として用いられている。
※2:TORC1
真核生物に広く保存されたプロテインキナーゼ複合体。Target-of-rapamycin protein kinase complex 1の略。栄養シグナリングのマスターレギュレーターとして知られる。細胞の成長や増殖、遺伝子発現、オートファジー、寿命など多くの生命現象に関与している。
※3:Greatwall
真核生物に広く保存されたプロテインキナーゼ。元々は、細胞分裂における染色体構造の維持に関与する因子として発見され、染色体を「護る」ことから名付けられた。細胞周期調節因子として知られる。
※4:PP2AB55δ
真核生物に広く保存されたプロテインフォスファターゼ複合体。Protein phosphatase 2A with a regulatory subunit B55δの略。PP2Aは、制御サブユニット(複合体を構成する単一分子)の違いによって機能が異なり、B55δ制御サブユニット(Saccharomyces cerevisiaeではCdc55p)を有するPP2AB55δは、細胞の成長や増殖、細胞周期などの調節因子として知られる。
※5:メタボローム解析
細胞内の代謝産物を一斉に解析する手法。
【解説図】
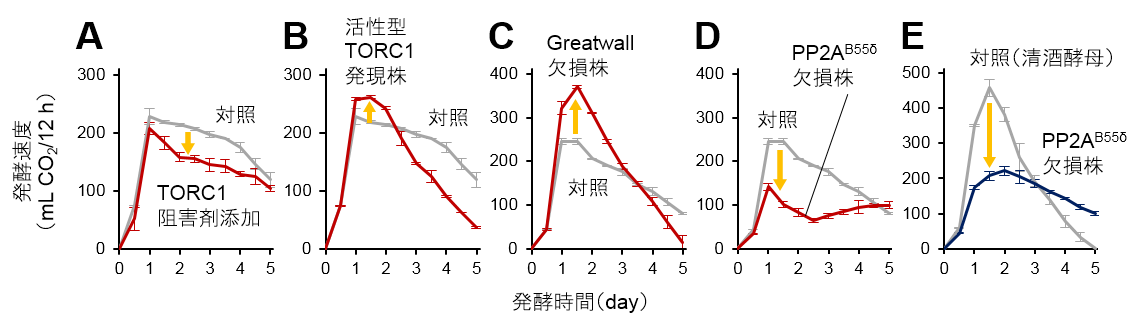
図1:TORC1-Greatwall-PP2AB55δ経路が発酵力に及ぼす影響
A: TORC1阻害剤(ラパマイシン)添加の影響、B: 活性型TORC1(TOR1L2134M)発現の影響、
C: Greatwall欠損(RIM15遺伝子破壊)の影響、D: PP2AB55δ欠損(CDC55遺伝子破壊)の影響、
E: 清酒酵母におけるPP2AB55δ欠損(CDC55遺伝子破壊)の影響
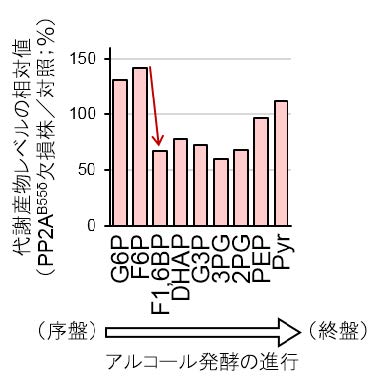
図2:PP2AB55δ欠損がアルコール発酵の中間代謝産物に及ぼす影響
グラフは、PP2AB55δ欠損(CDC55遺伝子破壊)株での各代謝産物のレベルを野生株との相対値で示す。矢印は、F6Pの蓄積とF1,6BPの減少を示しており、PP2AB55δ欠損によりPFK活性が低下したことを表している。

図3:発酵過程におけるTORC1活性
TORC1の直接の基質であるSch9pのリン酸化を、タンパク質混合物から単一の分子を分離するウェスタンブロット法により解析することでTORC1の活性を調べた。リン酸化型Sch9pのシグナルが強いほどTORC1活性が高いことを示す。
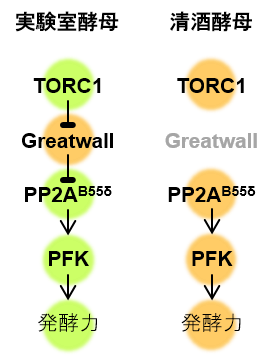
図4:TORC1-Greatwall-PP2AB55δ経路を介した発酵調節メカニズムのモデル
低活性をグリーン、高活性をオレンジで表す。清酒酵母は、TORC1活性が高くGreatwallが欠損していることからPP2AB55δの機能が強化されており、その結果高い発酵力を示すことが明らかになった。
【ストレス微生物科学研究室】
研究室紹介ホームページ:http://bsw3.naist.jp/courses/courses305.html
研究室ホームページ:http://bsw3.naist.jp/takagi/
(2018年10月29日掲載)
この記事をシェア ![]()