Sexual reproduction is a critical process to enhance genetic diversity and thereby promotes evolution of organisms. The most important step in sexual reproduction is the formation of gametes with opposite mating types (sexes), which usually show distinct morphology and motility (termed oogamy). In evolution, well recogized morphological distinction between male and female bodies is a secondary diversification to facilitate gamete formation and mating.
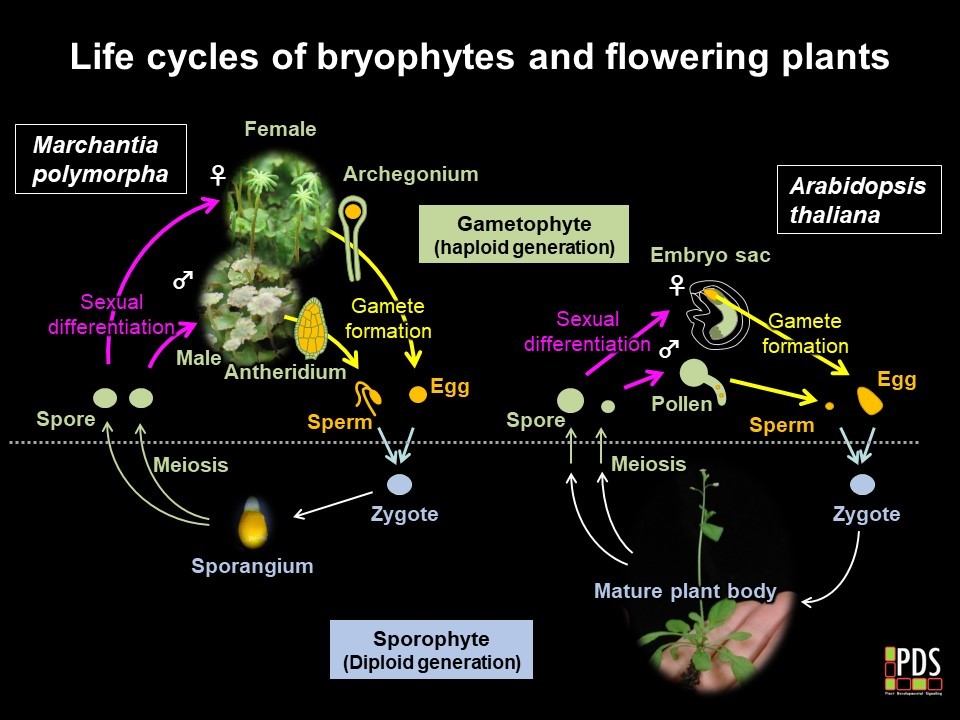
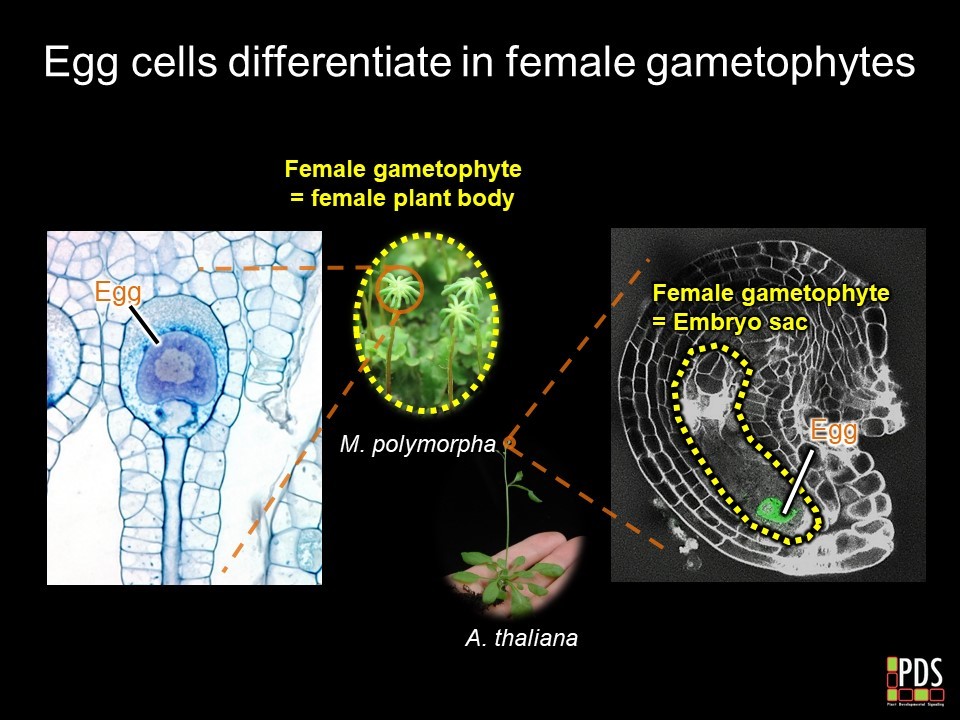
In animals, meiotic daughter cells directly differentiate into eggs and sperm. In plants, meiotic daughter cells divide further to produce multicellular haploid bodies called the gametophytes, in which male and female gametes differentiate. In angiosperms such as
Arabidopsis thaliana, male and female gametophytes are reduced to three-celled pollens and seven-celled embryo sacs, respectively. After fertilization, zygotes divide to produce multicellular diploid bodies called the sporophytes. Though this type of diploid-dominant life cycle is predominant, it limits the use of angiosperm models to investigate gamete differentiation in plants.
Bryophytes (mosses, liverworts and hornworts) retain ancestral haploid-dominant life cycles and thus can serve as a model systems to investigate sexual differentiation and gamete formation in plants. Among the bryophytes, the liverwort
Marchantia polymorpha is a superior model system, due to its well assembled genome sequnece with less redundant repertories of transcription factor genes. Availability of the asexual propagation route via gemma formation allows maintenance of gamete-deficient mutants and their phenotypic analyses.
Genes encoding RKD transcription factors are found ubiquitously in land plant genomes, but their biological functions have been enigmatic. In
A. thaliana, one
RKD genes is expressed in embryos and required for their development. Although, many other
RKD genes are expressed in egg cells, their roles in egg differentiation was difficult to prove in
A. thaliana. We identified a single
RKD gene in the
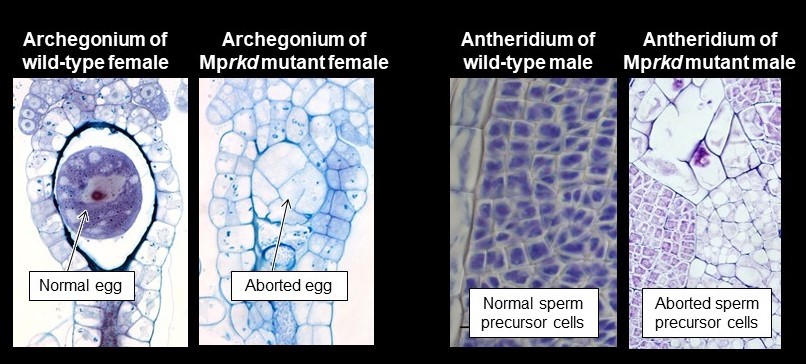
M. polymorpha genome and found that its knock-out mutants were speficically affected in egg and sperm maturation. Thus, by using
M. polymorpha, it became clear that the evolutionary origin of
RKD functions is to regulate gamete differentiation.
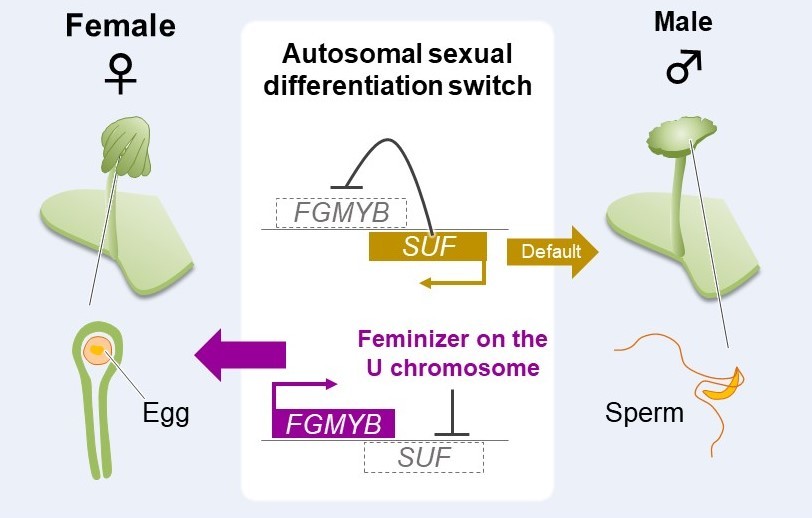
In another example, we found that a MYB transcription factor named FGMYB is strongly expressed in
M. polymorpha egg cells. Mutations in this gene resulted in female plants having male morphologies. In
A. thaliana, homologous
MYB genes were known to promote embryo sac differentiation. Because
M. polymorpha female bodies and
A. thaliana embryo sacs are evolutionarily equivalent, our study revealed that these MYB transcription factors are evolutionarily conserved regulators of female sexual differentiation in gametophytes.
The two studies mentioned above demonstrate the power of comparative molecular genetic studies of
M. polymorpha and
A thaliana in deciphering the mechanisms and evolution of sexual reproduction in land plants. We are currently investigating genes regulated by these transcription factors to understand genetic networks controlling sexual differentiation and their evolution in land plants.