
2023.06.01
種子から探る植物の細胞壁形成の分子メカニズム
植物代謝制御研究室・助教・國枝 正
- 要旨
- 細胞壁は植物細胞を象徴する構造体であり、セルロース微繊維やペクチンなどの多糖分子が相互作用し合いながら形成された複雑な構造をもっています。細胞の分化や成長、組織によって細胞壁はその構造を変化させるため、細胞壁の形成メカニズムにはいまだ不明な点が多く残されています。私たちは、シロイヌナズナの種皮表皮細胞がもつ“ムシレージ”と呼ばれる細胞壁の形成に、ユビキチンE3リガーゼのFLYING SAUCER 1(FLY1)とFLY2の両遺伝子が関与していることを明らかにしました。さらに、FLY2がペクチン合成の場であるゴルジ装置に局在していることを突き止め、これらのことからFLY2がゴルジ装置のタンパク質にユビキチン分子を付加することによって、細胞壁形成を制御している可能性が示唆されました。本稿では、前半に植物細胞壁やムシレージの構造や形成について概説し、後半にムシレージにおけるFLYの機能について紹介します。
1.はじめに:植物細胞壁
植物の細胞は細胞壁に覆われています。細胞壁が担う役割は多岐に渡り、細胞の形態維持や細胞間接着、機械刺激や微生物に対する物理的な障壁としての機能などが挙げられます。すなわち、周囲の環境に適応しながら逞しく生きる植物にとって細胞壁は重要な構造であり、その形成メカニズムを明らかにすることは植物の生存戦略を理解する上で欠かせません。植物細胞壁は、形成されるタイミングによって“一次細胞壁”と“二次細胞壁”に分類されます。個体を構成するすべての細胞がもっている一次細胞壁は新たな細胞が誕生する細胞分裂や細胞の成長の際に形成される細胞壁で、主骨格であるセルロース微繊維をヘミセルロースのキシログルカンが架橋し、それら多糖分子の間の空間を充填性多糖のペクチンが埋める構造をしています。それに対して二次細胞壁は、細胞の成長後に一次細胞壁の内側に形成される細胞壁であり、かつ二次細胞壁をもつ細胞は個体の中で限られています。セルロース微繊維が構造の主骨格であることは一次細胞壁と共通しているものの、二次細胞壁ではセルロース微繊維の架橋は主にヘミセルロースのキシランが担い、フェノール化合物のリグニンがセルロース微繊維とキシランの間の空間を充填しています。私たちはこのような細胞の分化や成長状態によって特徴的な構造を構築する細胞壁がどのような分子メカニズムで形成されるのかを明らかにするために、道管や種皮の細胞がもつ細胞壁に注目して研究を行っています。
2.種子を覆う細胞壁:ムシレージ
バジルやチア、シロイヌナズナなど一部の植物種の種子では、“ムシレージ”と呼ばれる二次細胞壁をその種皮に形成します。実験植物のシロイヌナズナでは受精によって胚発生が始まると、胚や胚乳を覆う珠皮において種皮を形成するための細胞の分化が開始されます。珠皮の表皮細胞では、ペクチンをはじめとする細胞壁を構成する多糖分子の合成が細胞内で活発となり、それらの多くは小胞輸送によって一次細胞壁と細胞膜の間の空間(ムシレージポケット)に分泌されて蓄積することで、ムシレージが形成されます(図1A)1,2)。しかしながら、この段階のムシレージは、未完成な状態です。種子が完熟して吸水すると、ムシレージは膨潤してその体積を大きく増加させ、ムシレージポケットに収まらなくなります。ムシレージポケットの容積の限界を超えると、ムシレージは一次細胞壁を物理的に破壊して種皮表皮細胞から放出され、種子の周りにカプセル状に展開することで完成に至ります(図1B)。種子を覆うペクチンに富んだムシレージは柔軟性や保水性に優れた性質をもっており、乾燥などの発芽時に種子を取り巻く環境から加わるストレスから胚を保護する役割を担っています3,4)。私たちはムシレージを構成する細胞壁成分の合成や修飾、細胞膜外への分泌輸送、および種皮からの放出を制御する因子をこれまでに同定し、ムシレージ形成の分子メカニズムを明らかにしてきました5,6,7)。
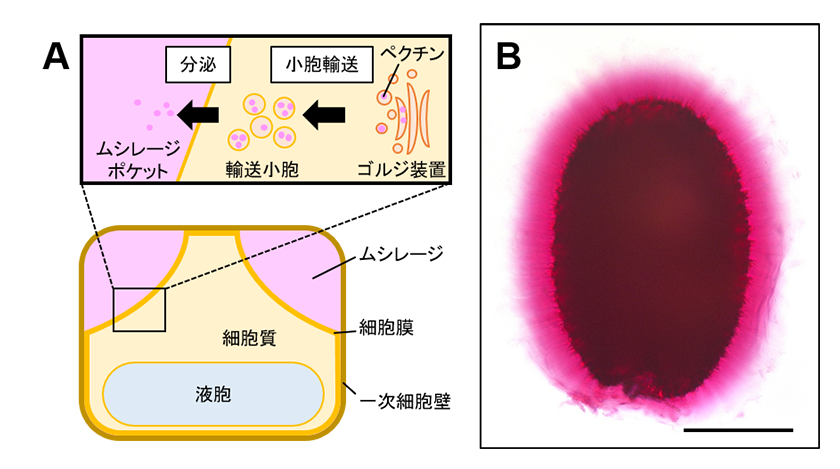
図1.
(A) ムシレージペクチンの合成後の細胞内輸送および蓄積モデル。
(B) シロイヌナズナ野生型種子のムシレージカプセル(赤色染色部)。スケールバー = 200 μm。
3.ペクチンのメチルエステル化修飾制御因子:FLYING SAUCER
2013年にカナダの研究グループから新規のムシレージ形成制御因子である“FLYING SAUCER 1(FLY1)”が報告されました8)。FLY1遺伝子を欠損する突然変異体(fly1)では、ペクチンのガラクツロン酸残基の脱メチルエステル化が亢進するために、ムシレージのカプセル形成異常を示します。FLY1は、ユビキチン分子を標的タンパク質に結合させる反応を促進する“ユビキチンE3リガーゼ”としての活性をもつことが示されています。しかしながら、膜貫通ドメインをもち、細胞内で機能するFLY1が、細胞膜外に存在するペクチンのメチルエステル化修飾をどのように制御しているのか、その分子メカニズムは現在までに明らかになっていません。
シロイヌナズナには、FLY1と相同性の高いユビキチンE3リガーゼであるFLY2が存在します。私たちはカナダの研究グループと協力して、FLY2の機能解析を行いました。まず、種子形成期のFLY2の遺伝子発現を検証したところ、発現量はFLY1と比較すると低いレベルであるものの、ペクチンが活発に合成される時期に珠皮表皮細胞で発現していることが分かりました。しかしながら、fly2変異体のムシレージに異常がないことは、先行研究によって示されています。そこで、fly1とfly2の二重変異体を作出して形成されるムシレージを観察しました。その結果、fly1 fly2二重変異体ではfly1単独変異体よりもムシレージのカプセル形成が顕著に阻害されることと、CaCl2処理によってfly1 fly2二重変異体のカプセル形成がさらに強く阻害されることが分かりました(図2)。Ca2+は脱メチルエステル化したペクチンのガラクツロン酸残基間でイオン架橋を形成することで、ペクチンゲルの物性を変化させることが知られています。これらの結果から、FLY2はFLY1とともにペクチンのガラクツロン酸残基のメチルエステル化を制御していることが示唆されました。
次に、膜タンパク質であるFLY2が細胞内で局在する部位を特定するため、蛍光タンパク質を利用した観察を行いました。分泌輸送に関わる主要な内膜系オルガネラとの共局在を検証したところ、FLY2はゴルジ装置と後期エンドソームに局在することが分かりました(図3)。ゴルジ装置がペクチンをはじめとする細胞壁の主要な構成成分の合成の場であることを考慮すると、FLY2がゴルジ装置においてペクチンのメチルエステル化修飾に関わっている可能性があります。以上の解析結果に加えて、陸上植物がもつFLY遺伝子の系統解析から、FLY1とFLY2はアブラナ科の出現とともにゲノム重複によって2遺伝子となったことを示唆する結果を得ています。このことは、遺伝子発現量を除いて、ムシレージ形成における両遺伝子の担う機能が等価であることを支持しています。
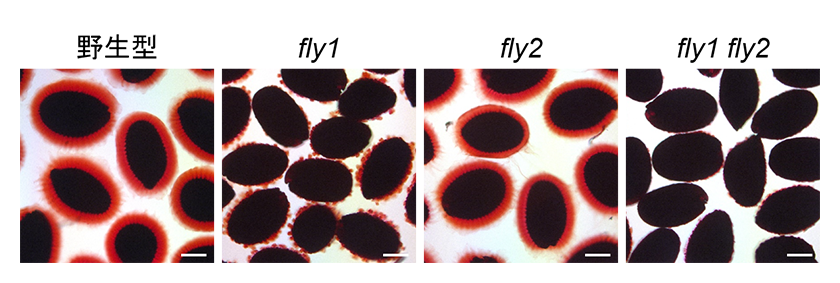
図2.野生型と各fly変異体の種子におけるCaCl2処理後のムシレージカプセル形成。スケールバー = 200 μm。
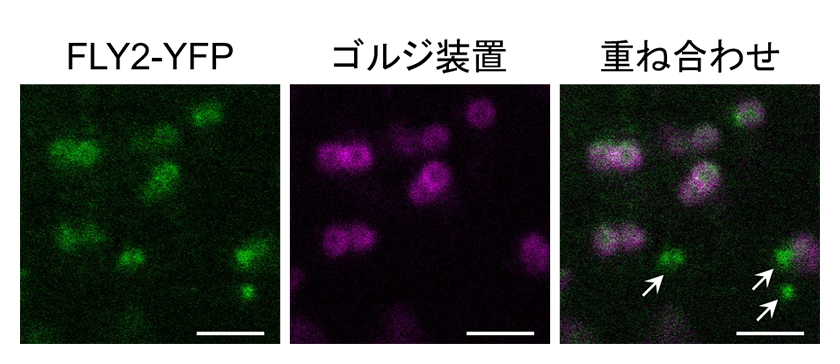
図3.FLY2に黄色蛍光タンパク質(YFP)を融合させたFLY2-YFPとゴルジ装置局在のタンパク質との共局在観察。ゴルジ装置と共局在しない一部FLY2-YFP(図中矢印)は、後期エンドソームに局在することが分かっている。スケールバー = 2 μm。
4.今後の展望
私たちの解析から、FLYはゴルジ装置で細胞壁ペクチンの修飾を制御していることが分かってきました。FLYはユビキチンE3リガーゼであることから、ゴルジ装置に局在するタンパク質のユビキチン化を介してペクチンのメチルエステル化を制御すると考えられます。しかしながら、そのユビキチン化標的タンパク質の実体は不明のままです。ペクチンは植物の細胞壁だけではなく、私たち人間の食にも広く利用されている有用物質です。今後は、FLYによってユビキチン化されるタンパク質を同定することで、ペクチンを中心に細胞壁形成の新たな分子メカニズムの解明につながることが期待されます。また、本稿では触れませんでしたが、FLY1とFLY2はいずれも分化中の道管細胞で発現することを見出しています。道管細胞も二次細胞壁を形成しますが、ムシレージとは物性が大きく異なって、リグニンに富んだ強固な細胞壁です。物性の異なる二次細胞壁の形成にFLYがどのように関わっているのか、そのメカニズムも興味深いです。細胞壁を対象にしながら研究に取り組むことで、植物が進化によって発達させてきたユニークな形態形成や生存戦略を明らかにしていきたいと思います。
参考文献
- Haughn, G. and Chaudhury, A. (2005) Trends Plant Sci 10, 472-477.
- North, H.M. et al. (2014) Ann Bot 14, 1251-1263.
- Penfield, S. et al. (2001) Plant Cell 13, 2777-2791.
- Rautengarten, C. et al. (2008) Plant J 54, 466-480.
- Kunieda, T. et al. (2013) Plant Cell 25, 1355-1367.
- Shimada, T. and Kunieda, T. et al. (2018) Plant Cell Physiol 59, 2331-2338.
- Takenaka, Y. et al. (2018) Nat Plants 4, 669-676.
- Voiniciuc, C. et al. (2013) Plant Cell 25, 944-959.
- Kunieda, T et al. (2020) Plant Cell Physiol 61, 308-317.

著者
國枝 正 Researchmap
略歴
2008年京都大学大学院理学研究科博士後期課程修了、博士(理学)
京都大学博士研究員、東北大学博士研究員、カナダUBC博士研究員、甲南大学特任研究助教を経て、2017年より現職。
- 研究内容:「細胞壁」、「膜交通」、「細胞小器官」、「種子」をキーワードに自生環境に巧みに適応して生きる植物の生存戦略を理解するための研究に従事。最近は、細胞壁の物性解析をきっかけにメカノバイオロジーの研究分野に挑戦しています。
國枝 正 NAIST Edge BIO, 0013. (2023)