成果報告
論文No.092
シロイヌナズナCGS1 mRNA分解ではポリA鎖の短縮化の停止が起こる
-
山下由衣、Ingrid Lambein、古林宗子、尾之内均、千葉由佳子、内藤哲
- Genes Genet. Syst. 88: 241-249 (2013)
Yamashita, Y., Lambein, I., Kobayashi, S., Onouchi, H., Chiba, Y. and Naito, S. (2013) A halt in poly(A) shortening during S-adenosyl-L-methionine-induced translation arrest in CGS1 mRNA of Arabidopsis thaliana. Genes Genet. Syst. 88: 241-249.
一般に真核生物におけるmRNA分解の最初の段階は、3'末端のポリA鎖の短縮化であり、この段階が律速段階と考えられています。本論文では、シロイヌナズナCGS1 mRNAの分解を誘導した条件下では、逆にポリA鎖の長いCGS1 mRNAが蓄積することを報告しました。
CGS1遺伝子はメチオニン生合成の鍵段階の酵素をコードし、その発現はメチオニンの代謝産物であるS-アデノシルメチオニン(SAM)に応答したリボソームの翻訳伸長の停止(翻訳アレスト)と、アレストしたリボソーム近傍でのCGS1 mRNA切断によってフィードバック制御されます。この制御におけるCGS1 mRNAのポリA鎖長の変化を調べた結果、通常条件下ではポリA鎖長は50-80塩基であったのに対し、分解を誘導した条件下では、ポリA鎖の長さが140-150塩基と10-30塩基の2つのCGS1 mRNA分子種が蓄積し、前者は切断前の全長mRNA、後者は切断後の分解中間体でした。
翻訳中のmRNAはキャップ結合タンパク質とポリA結合タンパク質の相互作用によって環状構造を取り、翻訳終結に伴ってポリA分解酵素がリクルートされると考えられています(図1)。翻訳アレストした状態ではポリA分解酵素がリクルートされず、ポリA鎖が長いままのCGS1 mRNA分子種が蓄積すると考えられます。そして、 CGS1 mRNA切断が起こるとポリA鎖が急速に分解されると考えられます。
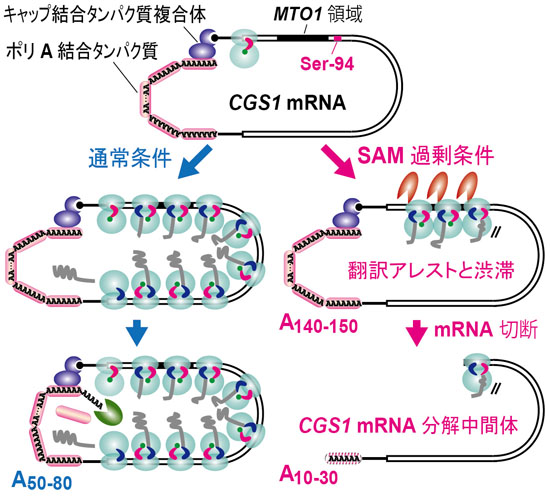
図1 翻訳中のmRNAはキャップ結合タンパク質複合体とポリA結合タンパク質を介して環状構造をとります。
左の経路:通常条件では、CGS1 mRNAは活発に翻訳されてポリソームを形成します。リボソームが翻訳終結すると、ポリA分解酵素(緑のPacman)が作用してポリA鎖は徐々に短縮化され、定常状態では約60塩基となります。
右の経路:SAM過剰の条件ではリボソームがSer-94コドンでアレストし、その後ろで渋滞を引き起こします。このとき、CGS1 mRNA上では翻訳終結が起こらないのでポリA鎖の短縮が抑制され、長いポリA鎖が保持されます。CGS1 mRNAの切断(赤のPacman)をきっかけに環状構造が解け、ポリA鎖は急速に短縮化されると考えられます。なお、MTO1領域はSAM応答に関わるシス配列です。