Research 研究成果の紹介
バイオサイエンス研究科ストレス微生物科学研究室の高木博史教授が公益財団法人アサヒグループ学術振興財団の平成25年度研究助成金対象者に選ばれました。
平成25年6月14日付けで、ストレス微生物科学研究室の高木博史教授が公益財団法人アサヒグループ学術振興財団の研究助成対象者に選ばれました。本助成は、主として食に関わる生活科学、生活文化および生活環境科学に関する研究を対象としており、高木教授は生活科学部門(食品や食料素材中に含まれる各種成分に関する基礎的・応用的研究や、食の機能性・安全性に関する解析・評価技術など、健康増進や生活改善に役立つことが期待される新規性の高い研究)に関する優れた研究として採択されました。
助成受託のコメント
アサヒグループ学術振興財団は、以前からビールを中心とする醸造科学の研究に助成 支援を行っていましたが、近年は食、生活、環境および医療に関わる研究に助成金を交付しています。私は18年前に食品企業の研究所から新設の公立大学に異動しました。その際、高等生物のモデルとして、また発酵産業において重要な酵母を用い、細胞が環境ストレスに適応する仕組みについての研究を学生と始めました。その結果、浸透圧調節物質として知られるプロリンが冷凍ストレスから酵母の細胞を保護することを見出しました。それ以来、プロリンの代謝制御機構と生理機能について解析し、得られた知見を産業酵母(パン、酒類、バイオエタノールなど)の育種に応用することを目指しています。今回頂いた助成金を有効活用し、ビール酵母の高機能化を目標に、さらに研究を発展させたいと思います。
支援を行っていましたが、近年は食、生活、環境および医療に関わる研究に助成金を交付しています。私は18年前に食品企業の研究所から新設の公立大学に異動しました。その際、高等生物のモデルとして、また発酵産業において重要な酵母を用い、細胞が環境ストレスに適応する仕組みについての研究を学生と始めました。その結果、浸透圧調節物質として知られるプロリンが冷凍ストレスから酵母の細胞を保護することを見出しました。それ以来、プロリンの代謝制御機構と生理機能について解析し、得られた知見を産業酵母(パン、酒類、バイオエタノールなど)の育種に応用することを目指しています。今回頂いた助成金を有効活用し、ビール酵母の高機能化を目標に、さらに研究を発展させたいと思います。
助成受託の説明
<研究テーマ>
プロリン代謝に着目したビール酵母の育種とビールの機能性向上への応用
<説明文>
アミノ酸のプロリン(Pro)には細胞やタンパク質を様々なストレスから保護する機能があります(図1)。ブドウ果実などに豊富に含まれ、酵母Saccharomyces cerevisiaeの主要なエネルギー源や窒素源となっており、ミトコンドリアのProオキシダーゼPut1により酸化されます(図2)。一方、ヒトでは栄養ストレス時に、細胞外マトリックス成分の分解とそれに伴うProの酸化がATP産生に寄与し、細胞の生存に寄与しています。また、植物や細菌では、乾燥・高浸透圧ストレス時にProを適合溶質として細胞内に蓄積しますが、ストレスからの回復時にはProの急激な分解とエネルギー産生を伴い、ストレス後の迅速なエネルギー供給源として機能します。このように、真核生物におけるProの分解はミトコンドリアで起こり、細胞の運動、生存、ストレス耐性など様々な生命現象に関わっています。この分解に際して、Proは細胞質ゾルからミトコンドリアに輸送されることが必要です。ミトコンドリアにはアミノ酸のトランスポーターが存在していますが、Pro特異的なトランスポーターについては、同定されていません。本研究では、真核生物のモデルとして、またビール産業において重要な酵母を用い、Proをミトコンドリアに輸送するトランスポーターを同定し、Pro代謝を介したエネルギー生産やレドックス制御への理解を深めます。また、トランスポーターの発現を人為的に制御したビール酵母を作製し、発酵生産における有用性を評価します。
<図の説明>
図1:プロリンの生理機能
プロリンは様々なストレスから細胞やタンパク質を保護し、図のような生理機能が考えられていますが、詳細な機構は明らかになっていません。また、植物や細菌では、乾燥・塩などのストレスに応答し、プロリンが細胞内に蓄積されます。一方、酵母ではストレスに曝されると、トレハロースやグリセロールを蓄積しますが、プロリン代謝関連遺伝子の転写量やプロリン含量に変化はありません。
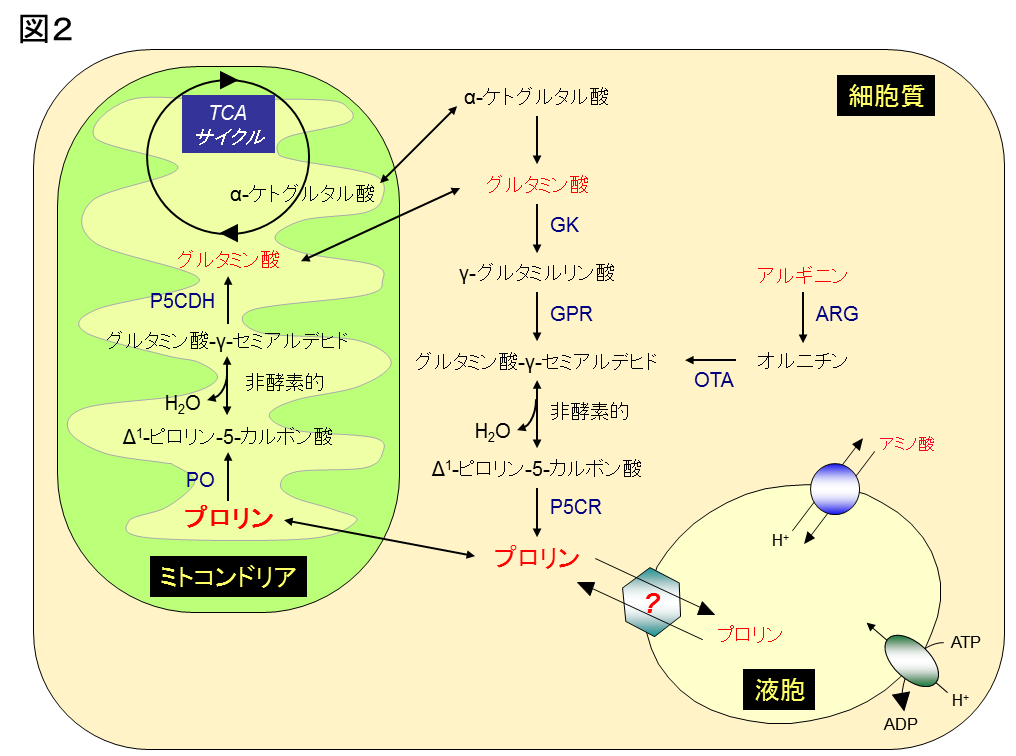
図2:酵母におけるプロリンの代謝経路
酵母では、プロリンは細胞質でおもにグルタミン酸から3種類の酵素(GK, GPR, P5CR)により合成されます。一部はアルギニンからアルギナーゼ(ARG)、オルニチントランスアミナーゼ(OTA)によっても合成されます。一方、過剰のプロリンはミトコンドリアで2種類の酵素(PO, P5CDH)により酸化的に分解され、グルタミン酸に変換されます。また、PO遺伝子の破壊により分解系を遮断すると、過剰のプロリンはおもに液胞に局在します。しかしながら、プロリンがミトコンドリアや液胞に輸送される分子機構は明らかになっていません。 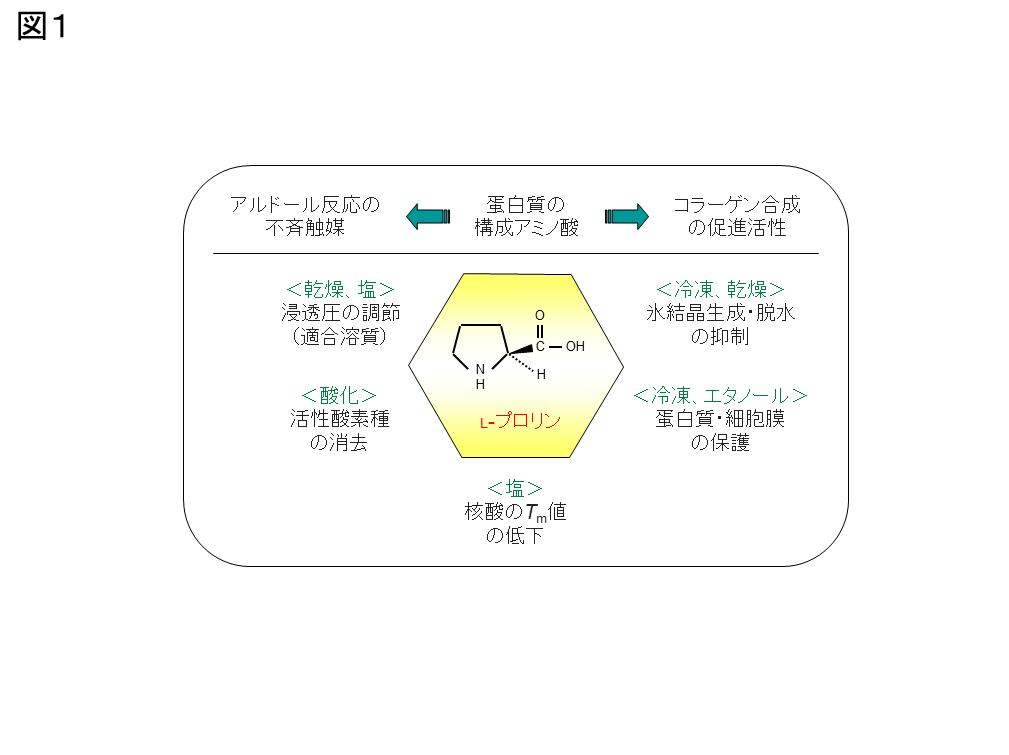
関連する論文・特許
- Y. Tatehashi and H. Takagi (2013) Characterization of γ-glutamyl kinase mutants from Saccharomyces cerevisiae. J. Biosci. Bioeng, DOI: 10.1016/j.jbiosc.2013.05.018.
- Y. Sasano, Y. Haitani, K. Hashida, I. Ohtsu, J. Shima and H. Takagi (2012) Enhancement of the proline and nitric oxide synthetic pathway improves fermentation ability under multiple baking-associated stress conditions in industrial baker's yeast. Microb. Cell Fact., 11:40 doi:10.1186/1475-2859-11-40.
- Y. Sasano, Y. Haitani, K. Hashida, I. Ohtsu, J. Shima and H. Takagi (2012) Simultaneous accumulation of proline and trehalose in industrial baker’s yeast enhances fermentation ability in frozen dough. J. Biosci. Bioeng., 113, 592-595.
- Y. Sasano, Y. Haitani, I. Ohtsu, J. Shima and H. Takagi (2012) Proline accumulation in baker’s yeast enhances high-sucrose stress tolerance and fermentation ability in sweet dough. Int. J. Food Microbiol., 152, 40-43.
- T. Kaino, T. Tateiwa, S. Mizukami-Murata, J. Shima and H. Takagi (2008) Self-cloning baker’s yeasts that accumulate proline enhance freeze tolerance in doughs. Appl. Environ. Microbiol., 74, 5845-5849.
- H. Takagi (2008) Proline as a stress protectant in yeast: physiological functions, metabolic regulations and biotechnological applications. Appl. Microbiol. Biotech., 81, 211-223.
- 高木博史 (2011) プロリン蓄積型形質転換酵母とその作出方法及び該酵母の利用方法. 特許第4837335号.
(2013年08月01日掲載)
この記事をシェア ![]()