成果報告
論文No.093
数珠つなぎになったリボソーム:翻訳停止したリボソームの後ろで渋滞したリボソームはどのような状態で停止しているのか
-
山下由衣、門倉嘉知、反田直之、藤原徹、瀧川一学、佐竹暁子、尾之内均、内藤哲
- J. Biol. Chem. 289: 12693-12704 (2014)
Yamashita, Y., Kadokura, Y., Sotta, N., Fujiwara, T., Takigawa, I, Satake, A., Onouchi, H. and Naito, S. (2014) Ribosome in a stacked array: Elucidation of the step in translation elongation at which they are stalled during S-adenosyl-L-methionine-induced translation arrest of CGS1 mRNA. J. Biol. Chem. 289: 12693-12704.
高等植物におけるメチオニン生合成の鍵段階を触媒するシスタチオニンγ-シンターゼをコードするCGS1遺伝子の発現は、メチオニンの代謝産物であるS-アデノシルメチオニン(SAM)に応答したSer-94コドンでの翻訳停止と、これと共役したmRNA分解によって調節されます。本論文では、翻訳停止にともなって渋滞した後続のリボソームがどのコドンで停止しているのか、また、渋滞したリボソームがどの段階で停止しているのかを調べました。翻訳伸長反応は、図1のように3つの段階からなります。既に、SAMに応答してSer-94コドンで停止したリボソームは転座前の段階で停止していることを明らかにしています。
本論文では追突した2つ目、3つ目のリボソームが、それぞれVal-85, Ala-76で停止している、つまり、リボソームが9コドン間隔で数珠つなぎになっていること明らかにしました。リボソームの停止位置とCGS1 mRNAの切断位置は非常によく対応しており、渋滞したリボソームに挟まれた位置でmRNAの切断が起こると考えられます(図2)。また、アミノアシル-tRNAのアナログであるピューロマイシンとの反応速度は、翻訳伸長の各段階で変化します。渋滞したリボソームのピューロマイシン反応を調べた結果、先頭のリボソームが転座前の段階で停止しているのに対して、追突したリボソームは転座途上の「ハイブリッド状態」で停止していると考えられました(図2)。本論文は、渋滞したリボソームの状態を明らかにした最初の報告です。
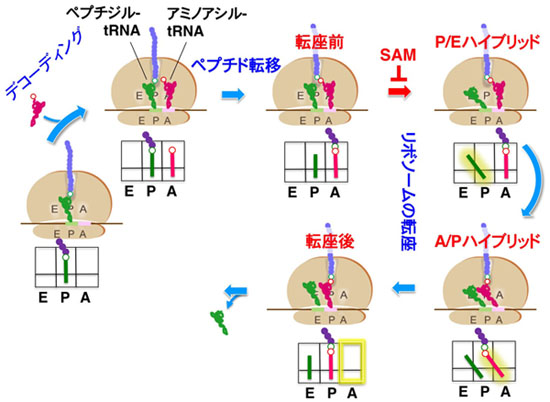
図1 リボソームにおける翻訳伸長反応
mRNAのコドンに対応するアミノ酸を付けたアミノアシル-tRNAがリボソームのA部位に入るデコーディング、P部位に位置する翻訳途上のペプチドを付けたペプチジル-tRNAとの間でペプチド結合を形成してA部位のtRNAにペプチドが転移するペプチド転移、リボソームが次のコドンに移る転座の各段階からなります。転座段階は、さらに大・小サブユニットでtRNAの位置が異なる「ハイブリッド状態」を経て進行します
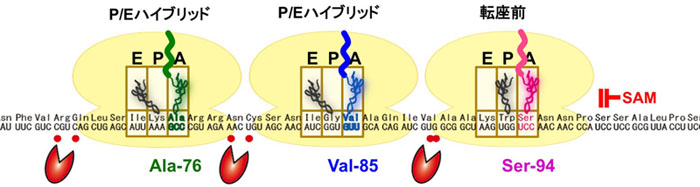
図2 SAMによって引き起こされるCGS1 mRNA上で渋滞したリボソームの状態
Ser-94で停止したリボソームの後ろに、9コドン間隔で後続のリボソームが数珠つなぎになります。先頭のリボソームの90%は転座前の段階で停止しており、渋滞したリボソームの大半は転座途上のハイブリッド状態で停止していると考えられます。CGS1 mRNAの分解中間体の5'末端位置(赤丸)は、渋滞したリボソームに挟まれた位置に対応しています。