Various biomolecules, such as proteins, RNA, and DNA, are involved in biological phenomena.
In order to elucidate the molecular mechanisms underlying the dynamic conformational changes of these biomolecules at the atomic level, we conduct fundamental research using structural biological analysis combined with new research methods/techniques.
Our laboratory provides an environment where students can devote themselves to their research, and we guide them to publish original papers backed by precise results.
Research activities require a variety of skills, from experimental planning to publication.
We hope our graduate students develop these abilities through their research activities.
Research Outline
The first step of our typical strategy is to elucidate the protein structure at the atomic and amino acid levels.
By obtaining detailed structural information of target proteins, much more insight into how these proteins can function.
Getting high-resolution structures is the most significant advantage of uncovering the details of protein structures.
The next step is to reveal proposed molecular mechanisms based on protein’s structural information by performing functional analyses.
Recently, we are also attempting to visualize protein dynamics by single-molecule analyses.
By utilizing several different methods for our research, our results provide new concepts that will change the contents of textbooks.
Research Highlights
Our research includes many fundamental/basic topics and can be summarized as "in-textbook-quality research."
We are studying various proteins, including drug target proteins, transport proteins, and motor proteins.
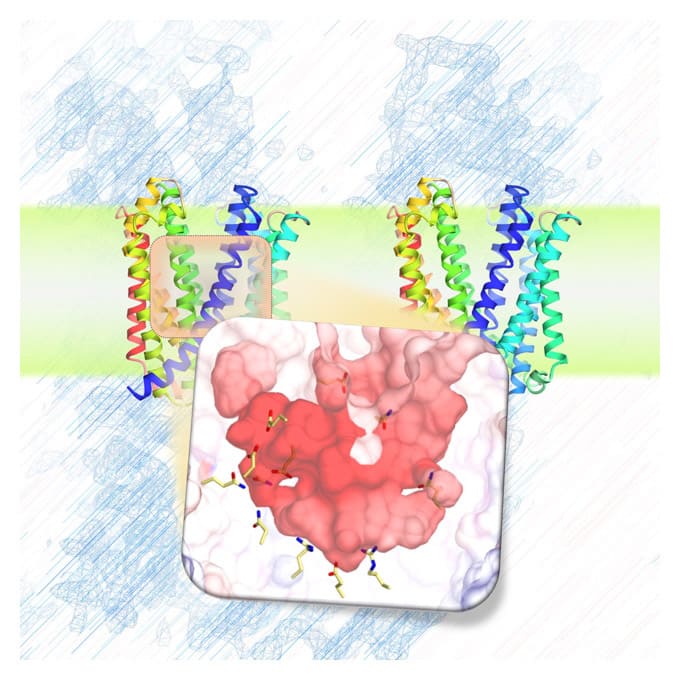
Atomic-level Visualization of the Molecular Mechanism of the Sec Membrane Protein Complex
The Sec membrane protein complex, Sec machinery, is located in the cytoplasmic membrane of bacteria and the endoplasmic reticulum membrane of eukaryotes and is an essential apparatus for the translocation of newly synthesized proteins across the membranes.
Many studies have been published on protein translocation, including the Nobel Prize-winning "signal hypothesis" proposed by Broebel et al. in 1975.
We have determined the detailed structure of each Sec protein and have been analyzing its function based on the structural information to elucidate the structural changes that occur during protein translocation across the membrane.
In addition, we are expanding our research to elucidate the overall structure of the Sec membrane protein complex at the atomic level, as well as trace time-dependent reactions to elucidate the dynamic mechanism.
Elucidation of Precise Mechanisms of Transporters and Channels
Biological membranes separate cells from the outside world, as well as organelles within cells.
Many membrane proteins are involved in the uptake and efflux of substances, such as drugs and ions, as well as information transmission and energy synthesis.
Therefore, detailed three-dimensional structures of membrane proteins are required for elucidating their molecular mechanisms, and they provide essential information for understanding biological phenomena.
Analysis Using the Latest Technology
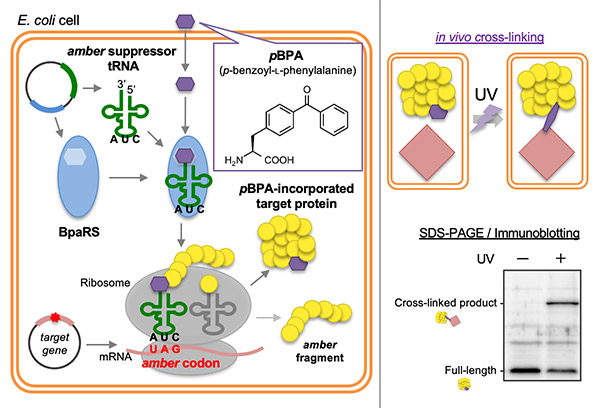
In order to track the transportation/movement of proteins, we analyze the protein dynamics in vivo using the PiXie (pulse-chase and in vivo photo-cross-linking experiment) method, and observe one-unit dynamics using high-speed atomic force microscopy and high-performance cryo-electron microscopy through collaborative research, and perform other analyses using the latest technologies.
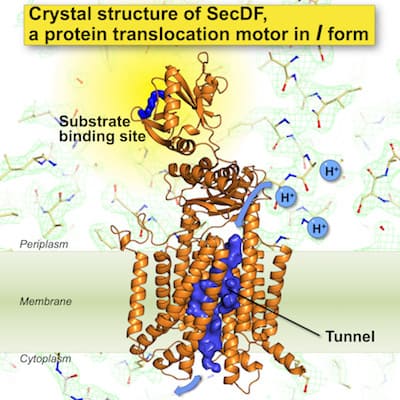
The crystal structures of SecDF were determined at high resolution.
The structures suggested the presence of a site interacting with protein substrates and a pathway for proton ion.
Knowledge of the transmembrane protein export mechanism will provide a structural basis for the development of new type of antibiotics.
Summary
The cell membrane separates the cell’s interior from the environment. Extracellular transport of proteins synthesized in the cytoplasm is an essential process for sustaining life. We conduct basic research on the molecular mechanisms of the protein transport.
In bacteria such as Escherichia coli, the membrane-embedded SecDF is involved in protein transport. Specifically, SecDF mediates the translocation of its substrates by using the H+ gradient across the membrane. However, the detailed molecular mechanisms of SecDF remains unclear. In particular, the sites that interact with substrate proteins passing through the membrane and the H+ pathway remained elusive. Recently, we successfully determined the crystal structure of SecDF at high resolution (2.6 Å). Based on this structural information, we carried out biochemical experiments and computational molecular dynamics simulation. The results of these analyses identified SecDF’s a substrate-binding site and a H+ ion pathway. Moreover, we demonstrated that the each SecDF domain undergoes dynamic conformational changes, driven by the H+ ion gradient. Based on these findings, we proposed a novel molecular mechanism for SecDF-driven protein translocation across the membrane. Our findings regarding this essential biological phenomenon will contribute to progress in basic life science. In addition, our findings will provide a theoretical basis for developing new antibiotics that target the bacteria-specific SecDF protein.
Study Description
(A) Background
Cellular compartments are distinctly separated from their exterior environment by the membrane. Although the cell membrane does not allow free passage of molecules and ions, their bidirectional transport across the membrane is necessary for maintaining life. Proteins, which are synthesized by cytoplasmic ribosomes, are necessary components of living organisms. Approximately 30% of proteins are translocated across the cell membrane to perform various activities in the extracellular milieu. Protein translocation is mediated by the ubiquitous Sec translocon channel. Because the Sec translocon functions as a passive channel, accessory proteins must assist in protein secretion. In bacteria, the motor proteins SecA and SecDF drive the polypeptide translocation. SecDF is a bacteria-specific membrane protein consisting of transmembrane and periplasmic domains. It is widely believed that along with the influx of H+ through the transmembrane domain, SecDF undergoes repeated cycles of conformational transition from the F-form (capturing state) to the I-form (holding state) in order to undergo translocation of polypeptides released from the Sec translocon to the periplasm. However, previous studies reported only the full-length F-form of the SecDF structure, underscoring the need for high-resolution structure of SecDF in I-form .
(B) New Findings
We successfully determined the crystal structures of I-form SecDF from Deinococcus radiodurans, at a resolution of 2.6 Å. The lipidic cubic phase (LCP) crystallization technique was applied to crystallize SecDF, and X-ray diffraction data sets were collected at beam line BL32XU at SPring-8, Hyogo, Japan. The I-form SecDF structure exhibits two noteworthy characteristics. First, the cavity of the P1 domain, the first periplasmic domain, was occupied by a small molecule, probably mimicking the binding of substrate protein. We conducted a biochemical study to identify the interaction site in SecDF with the protein substrate, and found that a substrate-specific interaction site is located in this cavity. These results strongly suggest that the cavity forms the substrate-binding site. Second, the transmembrane region of SecDF possesses a tunnel architecture that spans across the membrane form the cytoplasm to periplasm. This tunnel seems to be the H+ ion pathway, because an aspartate residue essential for SecDF functions (i.e., proton transfer and preprotein translocation activation) was located at the center of the tunnel. We performed molecular dynamics simulations of SecDF; the results implied that the deprotonated and protonated states of the aspartate residue are closely related to the opening and closing of the tunnel, respectively, and that water molecules are aligned via hydrogen bonding along the inner surface of the tunnel ranging between its cytoplasmic and periplasmic ends. These results suggest that the tunnel allows for H+ ion entry into the cytoplasm from the periplasm. Moreover, we found that H+ ion influx and preprotein translocation are prohibited when the highly flexible P1 domain is rigidly fixed. This finding suggested an association between P1 domain flexibility and H+ ion conductance activity. Based on these findings, we proposed a new working model for SecDF: i) the F-form conformation of SecDF captures the substrate protein in the P1 cavity; ii) SecDF undergoes the conformational transition from the F- to the I-form with the substrate. the conformational F-to-I transition is driven by energy derived from the H+ ion influx, facilitated by water molecules in the transmembrane domain tunnel; and iii) preprotein translocation is achieved by repeated cycles of the conformational F-to-I transition.
(C) Significance and Future Perspectives
In this study, we revealed the molecular mechanisms of SecDF motor. Elucidation of the protein translocation mechanisms, which are essential for life, will greatly contribute to basic research in related fields. Because SecDF is specific to and essential for bacterial survival, knowledge of SecDF mechanisms will provide a technical framework for the discovery of new antibacterial agents.
High-resolution structural and functional analysis of SecYEG
A proposed new mechanism for protein translocation across the membrane: Opening and closing mechanism of the cap of the protein-conducting channel
The structure of the protein-conducting channel, an essential machinery for all organisms, was determined at the highest resolution.
A new mechanical model was proposed for protein translocation across the membrane: the protein-conducting channel possesses a cap that regulates the translocation of proteins.
The findings will help advance basic research on protein translocation across the membrane.
Summary
One of the most basic biological processes observed in all organisms is transport of newly synthesized proteins across the cell membrane. This process is necessary for delivery of proteins to the locations where they function. To serve that purpose, the biomembrane contains a machinery known as the protein-conducting channel (Sec translocon). Several structural studies showed that the Sec translocon forms several transition states, and several protein translocation mechanisms have been proposed. However, the paucity of available data prevented in-depth discussion of the mechanisms and warranted high-resolution analyses of the Sec translocon structure.
We determined the structure of the bacterial SecYEG translocon complex at the highest resolution (2.7 Å resolution). Based on the structural information, we performed functional analysis, which suggested that in the resting state, the channel is covered by the cytoplasmic loop of the complex subunit SecG, and that the SecG cap is removed when protein translocation starts. Our study provided a reliable theoretical model for Sec translocon–mediated protein translocation.
Study Description
(A) Background
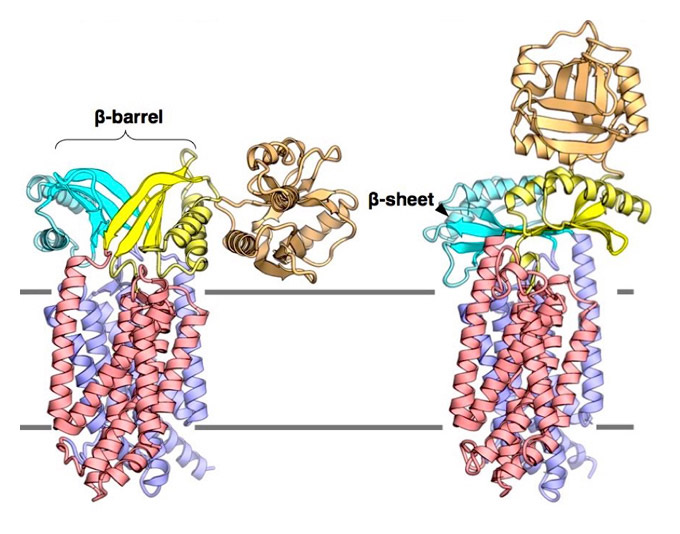
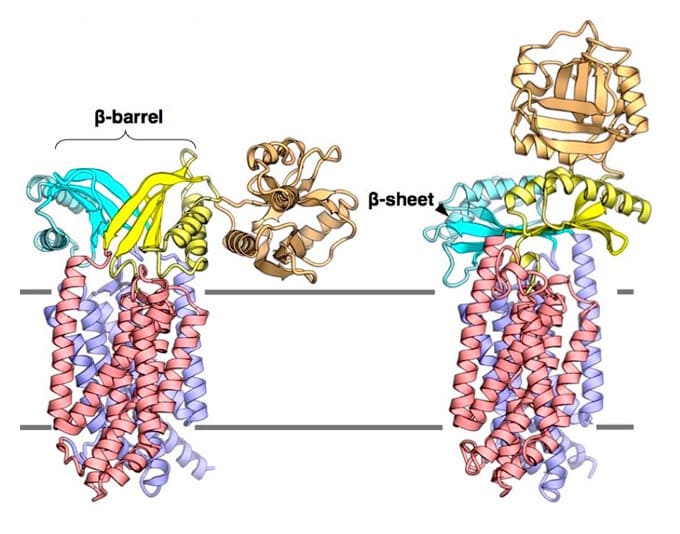
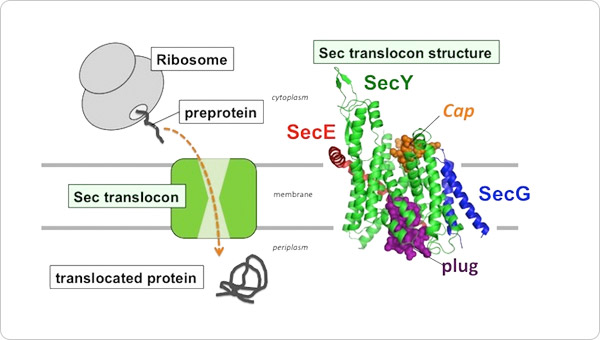
The cell contains different membrane-separated compartments. The cell needs to transport material and signals appropriately across those biomembranes to maintain life. In the cytoplasm, ribosomes synthesize a wide variety of proteins. Newly synthesized proteins are transported to the appropriate locations that require their functions and they fold into their functional shapes. To this end, proteins must pass through the biological membrane via a channel called the Sec translocon complex (SecYEG in prokaryotes and Sec61αγβ in eukaryotes, see Figure 1, left panel).
A great deal of research effort has been devoted to elucidating how the translocon allows for the passage of large protein molecules while maintaining the permeability barrier. Previously, several Sec translocons were structurally analyzed, and those studies provided a basis for proposing models of transmembrane protein translocation. However, further discussion of the mechanisms required more precise information about the Sec translocon structure.
Figure 1. Left panel: Schematics illustrating protein translocation across the membrane.
The figure illustrates the process by which the substrate protein is transported to the extracellular space via the Sec translocon. Right panel: A ribbon presentation of the SecYEG structure determined at 2.7-Å resolution. The SecY, SecE, and SecG subunits are indicated in green, red, and blue, respectively, whereas the cap and plug regions are indicated in orange and purple, respectively. The illustration emphasizes the visibility of the plug and cap regions for reference. SecG is located peripheral to SecY, and the SecG loop (cap) is positioned to seal the SecY channel.
(B) New Findings
We successfully purified SecYEG from Thermus thermophilus, a species of highly heat-resistant bacteria, and determined its crystal structure at of 2.7 Å resolution .The lipidic cubic phase (LCP) crystallization technique was employed to crystallize SecYEG, and X-ray diffraction data sets were collected at the BL32XU beam line at SPring-8, Hyogo, Japan.
In previous crystallographic studies on the SecYEG complex, 4.5 Å was the highest resolution achieved. The 2.7-Å resolution achieved in this study allowed more precise structural analysis. Notably, this high-resolution structure revealed the presence of hitherto-unknown loops that link different domains, allowing us to develop a precise structural model that specifies the locations of almost all amino-acid residues (Figure 1, right panel). The new findings regarding the SecG loop are of major importance. In particular, the SecG loop covers the channel of SecY. To investigate the role of the SecG loop, we conducted an in vitro protein translocation assay using genetically engineered SecYEG mutants, in which the SecG loop was disulfide-bonded to the cytoplasmic side of the SecY. These mutants did not exhibit translocation activity, although activity was restored after the bond was broken by addition of a reductant.
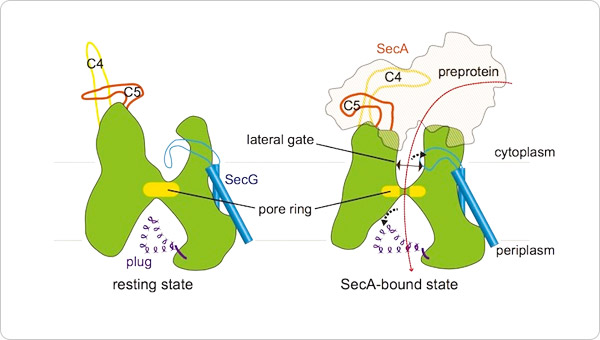
Based on these functional analyses, comparisons with previously reported molecular structures, and molecular dynamics simulations, we showed that (i) in the resting state of SecYEG, the SecG loop serves as a cap that seals the channel to prevent leakage of substances; and (ii) the cap is removed from the channel during translocation.
Previous work showed that the channel is sealed with a flexible plug (shown in purple in Figures 1 and 2) on the periplasmic side. Taking note of these findings, we proposed that in the resting state of the translocon, access to the channel is sterically hindered on both sides of the cell membrane, and the translocon undergoes conformational rearrangement upon receiving a protein translocation signal (Figure 2). In our original article, we also presented the crystal structures of the SecYEG translocon in other states. Specifically, we described the interactions between SecYEG and its substrate proteins. We also discussed the conformational transitions leading to opening of the lateral gate of the Sec translocon.
Figure 2. A cross-sectional model of SecY, illustrating protein transport through the channel.
In the resting state of SecYEG, the protein-conducting channel is sealed with a cap on its cytoplasmic side and a plug on its periplasmic side. During protein translocation, both the cap and plug moieties move out of the cavity to make room for protein transport (indicated by the arrow in the right panel).
(C) Significance and Future Perspectives
This study is the first to report the long-awaited high-resolution structure of the complete SecYEG translocon complex. The Sec translocon alone is not sufficient for protein translocation, which requires the engagement of other factors. In recent years, a great deal of effort has been devoted to electron microscopic studies of the ribosome-SecYEG complex. The structural model proposed in our study will aid in analyzing the electron microscopy images emerging from other work. Our study will help advance future basic research on protein transport, one of the processes essential for maintaining life. We expect that our results will inform future studies of the structure, function, and dynamics of the Sec translocon.
Structural analysis and function of the membrane protein insertase YidC
Kumazaki K, Chiba S, Takemoto M, Furukawa A, Nishiyama K, Sugano Y, Mori T, Dohmae N, Hirata K, Nakada-Nakura Y, Maturana AD, Tanaka Y, Mori H, Sugita Y, Arisaka F, Ito K, Ishitani R, *Tsukazaki T and *Nureki O. structural basis of sec-independent membrane protein insertion by YidC.nature 509,5120(2014).[naistar]
Highlights
The three-dimensional structure of the membrane protein insertase YidC was determined for the first time.
The mechanism of membrane protein insertion, a biological process conserved from bacteria to humans, was elucidated.
The results of this study will help advance life science research and provide a basis for developing new drugs.
Summary
The membrane protein insertase YidC is present in cell membranes of a variety of species and plays an indispensable role in maintaining cell viability. Cells are the smallest unit of life, and knowledge of cell membrane function is essential for understanding the fundamental life processes common to microbes and humans. However, despite extensive research, the molecular structure of YidC remained unknown for a long time, and the molecular mechanism involved in incorporating proteins into the membrane was unclear. We determined the first crystal structure of YidC. The structure reveals that YidC has a novel structure that contains a hydrophilic (or “water-loving”) cavity within the hydrophobic (or “water-repelling”) environment of the membrane. We showed that the proteins inserted into the cell membrane interact with YidC’s hydrophilic pocket. These findings led to a new model for YidC-mediated membrane protein insertion. Our study elucidated a fundamental biological process conserved from bacteria to humans. Because YidC is essential for bacterial growth and survival, the findings of this study will contribute to future life science research, providing a basis for developing new antibiotic agents that target the YidC proteins of pathogenic microorganisms.
Study Description
(A) Background
The plasma membrane, which consists of two layers of lipid molecules, contains a large number of proteins that mediate a wide array of biological processes. Membrane-segregated cells are the fundamental unit of life. Cells have organelles, which are also surrounded by biomembranes. Organelles play vital roles in maintaining and regulating cellular functions. Biomembranes serve to separate the interior space from its exterior, functioning as a barrier. Biomembranes, including membrane proteins, play important roles for molecular transport, signal transmission, and energy production. Ensuring proper insertion of proteins into the biomembrane is essential for life. In a wide range of species, YidC is one of the membrane protein insertion machineries. This protein interacts with newly synthesized membrane proteins from ribosomes [Note 1] and incorporates them into the cell membrane. During this incorporation process, the hydrophilic extracellular domain of the membrane protein passes across the cell surface, while the hydrophobic transmembrane moiety remains within the cell membrane. In order to understand the basic principles of life, it is important to clarify the molecular mechanism by which YidC inserts membrane proteins. However, the three-dimensional structure of YidC remained unknown for a long time, and the mechanism of membrane protein insertion remained to be elucidated.
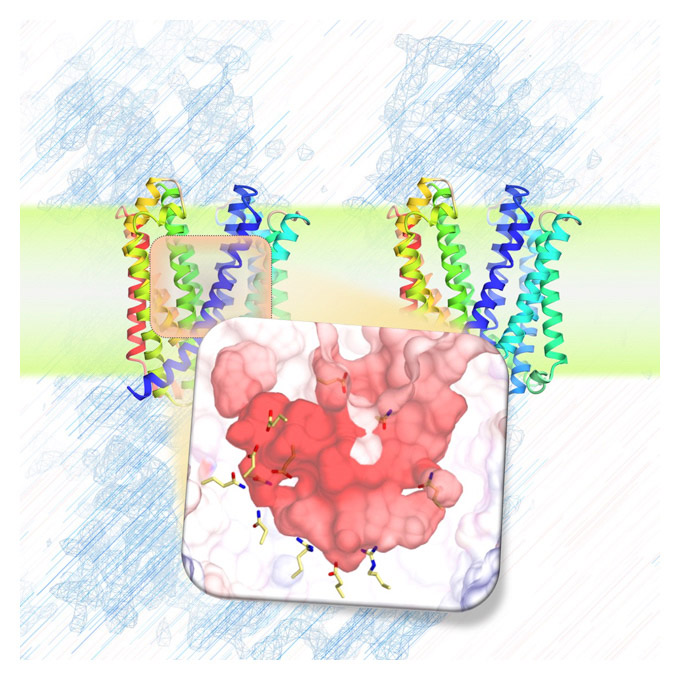
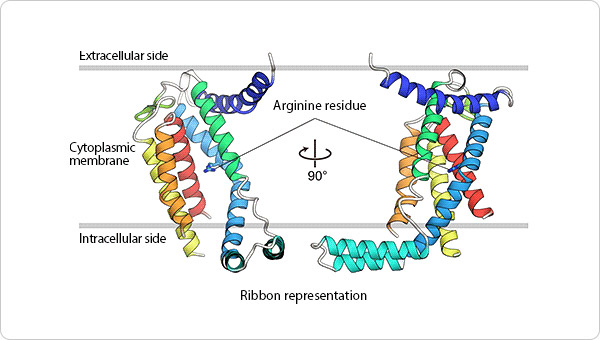
Figure 1. Detailed YidC structure.
The ribbon diagram schematic drawing depicts the YidC structure. The main structural components of YidC are five transmembrane alpha-helices (represented as ribbons), which are mostly embedded in the cell membrane. The position of the arginine residue, which plays an important role in YidC activity, is indicated by a stick model.
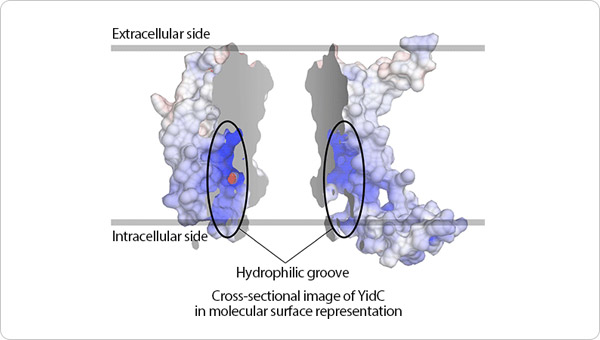
Figure 2. Molecular surface model of YidC.
In the model, positively and negatively charged moieties are indicated in blue and red, respectively. The protein is divided into two parts separated by a cross-section. The diagram shows that YidC possesses a positively charged hydrophilic pocket.
(B) Our Findings
We successfully determined the crystal structure of YidC derived from Bacillus halodurans [Note 2]. The lipidic cubic phase (LCP) crystallization technique was employed to obtain YidC crystals [Note 3]. X-ray diffraction data sets were collected at the BL32XU beam line at SPring-8, Hyogo, Japan [Note 4]. As a result, we successfully determined the YidC structure at a relatively high resolution of 2.4 Å (Figure 1). The three-dimensional structure of YidC revealed a large groove, which is positively charged and consists of hydrophilic amino acid residues (Figure 2). Unexpectedly, the hydrophilic groove of YidC is positioned in the hydrophobic milieu of the cell membrane. Based on the crystal structure of YidC, we carried out in vivo photo-crosslinking experiments [Note 5] and genetic studies using B. subtilis [Note 6]. These results led us to conclude that the hydrophilic YidC groove interacts with the membrane proteins (substrate). Among the hydrophilic amino acids that define the groove, the highly conserved arginine at position 72, a positively charged residue, is located at the center of the groove. We found that this amino acid residue is critical for the membrane protein insertion function of YidC. These and previous findings led us to postulate that membrane protein insertion takes place as the hydrophilic binding pocket of YidC recruits the extracellular (hydrophilic) segment of the substrate into the low-dielectric environment of the membrane (Figure 3). This study represents an important milestone in the study of the mechanism of membrane protein insertion.
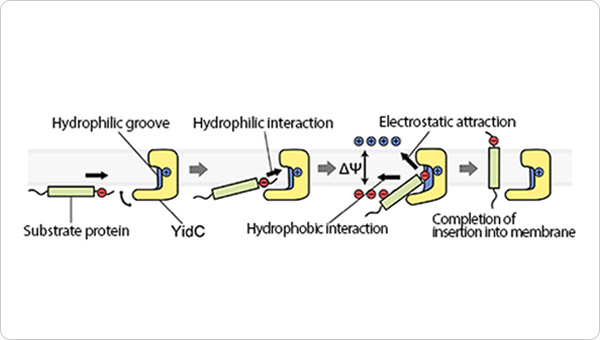
Figure 3. Schematic presentation of YidC-mediated membrane protein insertion.
(1) The positively charged YidC groove interacts with the negatively charged extracellular region of the substrate protein. (2) Insertion of the substrate molecule into the membrane layer is accelerated by hydrophobic interactions between the lipid chains and the hydrophobic intramembrane domain of the substrate, and by the electrostatic attraction of the positively charged extracellular component toward the extracellular space due to the membrane potential (difference in electrical potential across the membrane). (3) The substrate protein is released from YidC and becomes anchored in the cell.
(C) Significance and Future Perspectives
This structural biology study was the first to explain in detail how YidC mediates membrane protein insertion, a fundamental biological process observed in species from bacteria to humans. The findings of this study will contribute to basic research on protein production and transport. In addition, because YidC is essential to bacterial growth and survival, the findings of this study provide a basis for developing new antibiotic agents that target the YidC proteins of pathogenic microorganisms.
Terminology and Explanation
[Note 1: Ribosome]
The ribosome is the machinery that synthesize proteins based on genetic information in the cell. Due to its importance, it is a longstanding subject of research. The Nobel Prize in Chemistry 2009 was awarded for studies of the structure and function of the ribosome.
[Note 2: Crystal structure determination]
In this study, the protein structure was solved using X-ray crystallography techniques. In X-ray crystallography, the target material is irradiated with X-rays, and diffraction data are collected and analyzed to determine its structure.
This technique helps crystallize proteins reconstituted in lipid bilayers, and has been frequently used for crystallization of membrane proteins.
[Note 4: SPring-8]
One of the world’s largest synchrotron radiation facilities, located in Sayo-cho, Hyogo, Japan. This facility enables us to use powerful X-rays for irradiation experiments.
[Note 5: In vivo photo-crosslinking]
This method helps investigate the interactions between proteins in vivo by crosslinking them. In this study, this method was used to identify the interactions between the cavity of YidC and a YidC-dependent substrate.
[Note 6: Genetic study using B. subtilis]
In this study, by genetically modifying the chromosomal genes of B. subtilis, we created yidc mutant strains whose colony colors indicated different YidC activities. Using these mutants, we systematically investigated the activities of partially modified YidC mutants.
Highlights