成果報告
論文No.041
非典型的なNa+輸送活性を示すイネのOsHKT2;4輸送体によるK+輸送とMg2+, Ca2+イオンとの競合。
-
堀江智明、Brodsky, D. E.、 Costa, A.、 金子智之、Lo Schiavo, F.、且原真木、Schroeder, J. I.
- Plant Physiol. 156, 1493-1507 (2011)
Horie, T., Brodsky, D. E., Costa, A., Kaneko, T., Lo Schiavo, F., Katsuhara, M. and Schroeder, J. I. Plant Physiol. 156: 1493-1507 (2011)
植物のHKT型輸送体の内、クラスIIに分類されるグループは、Na+-K+共輸送活性を示し、かつ高濃度のNa+の存在下においては、Na+選択的輸送体として機能する事が知られている。
ジャポニカ米の栽培種日本晴には、2つの機能未知のクラスII輸送体、OsHKT2;3、OsHKT2;4が存在する。両者は、アミノ酸配列レベルで93%以上一致するにも関わらず、OsHKT2;3は、酵母やアフリカツメガエル卵母細胞(oocytes)内で顕著なイオン輸送活性を示さなかった。それに対し、OsHKT2;4は、両発現系において顕著なK+輸送活性を示した。興味深い事に、HKT輸送体の特徴である、高濃度のNa+の存在下におけるNa+選択的輸送活性は、OsHKT2;4では、著しく小さい事が判明した。
以前の報告から、OsHKT2;4は、一価のアルカリ陽イオンのみならず、Ca2+やMg2+を含む広範なイオン選択性を示し、特にCa2+の恒常性に対する重要性が示唆されていた。OsHKT2;4のCa2+やMg2+の透過性とK+の選択性との関連を評価するために、oocytesを利用した競合実験を行った。その結果、OsHKT2;4は、イオン種競合環境下においてK+に対する顕著に高い選択性を示す事が示された。これらの結果は、OsHKT2;4が、実際のイネ日本晴の中で、K+の恒常性により強く関与する生理機能を持つ輸送体である事を示唆している。
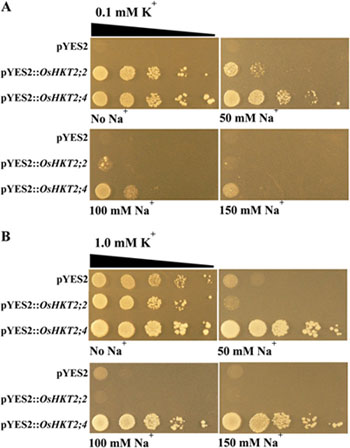
図1 OsHKT2;4依存のK+輸送欠損相補能は、他の代表的なイネのクラスII輸送体OsHKT2;2に媒介された場合よりも、高濃度のNa+による阻害効果が著しく低い。各コンストラクトで形質転換した変異酵母CY162細胞を、異なる濃度のK+, Na+を含むAP培地上で生育させた際の状況を示している。 A. 0.1 mM KCl、及び各濃度のNaClを含んだAP培地上で、各CY162細胞を生育させた際の写真。 B. 1.0 mM KCl、及び各濃度のNaClを含んだAP培地上で、各CY162細胞を生育させた際の写真。SC-Ura + 50 mM KCl培地で前培養された各形質転換体は、OD600を0.1に調整した後AP培地上にスポットされた(右へ移行するたびに10倍希釈されている)。全てのプレートを30℃で6日間静置した後、写真撮影した。
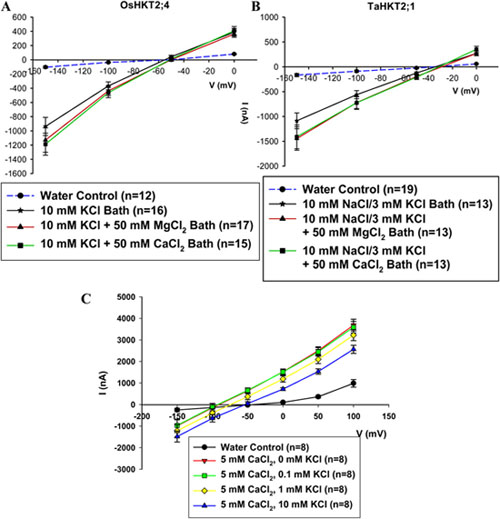
図2 アフリカツメガエル卵母細胞を用いた電気生理実験。K+, Na+イオン存在下における、OsHKT2;4およびTaHKT2;1のMg2+, Ca2+透過性の競合的阻害実験の結果を示す。OsHKT2;4に関しては10 mM K+存在下で、TaHKT2;1においては、10 mM K+, 3 mM Na+の存在下で、それぞれMg2+, Ca2+の透過性を解析した。A. 10 mM KCl溶液中に 50 mM MgCl2を添加する事によって、OsHKT2;4依存の逆転電位は、5 mV程正方向へ移行した。50 mM CaCl2の添加によっても、同様に僅か4 mV程度の逆転電位の正方向へ移行が観察された。B. 10 mM K+, 3 mM Na+溶液中に 50 mM MgCl2を添加する事によって、TaHKT2;1依存の逆転電位は、6 mV程正方向へ移行し、50 mM CaCl2の添加の場合は、その移行は、正方向へ3 mV程度に過ぎなかった。C. 5 mM CaCl2の存在下で、溶液中のK+濃度を0.1 mMから, 1 mM, 10 mMへと上昇させた場合、OsHKT2;4依存の逆転電位は、それぞれ正方向へ16 mV, 36 mVの移行を示した。エラーバーは、標準誤差を示し、実験条件に応じn=12-19の卵母細胞を使用した。